- Volume 60 , Number 3
- Page: 470–6
The significance of proteins actively secreted by Mycobacterium tuberculosis in relation to immunity and complications of mycobacterial diseases
It is widely recognized that Mycobacterium leprae is virtually nontoxic. Large amounts of bacilli may reside in the tissues of the infected host almost without clinical symptoms as in early lepromatous leprosy. Most symptoms and important complications of leprosy are due to immune reactions against antigens released from the bacilli. Nerve damage induced during reversal reactions is caused by delayed-type hypersensitivity (DTH) reactions against bacillary antigens released from Schwann cells, and the nerve is affected as an innocent bystander. Erythema nodosum leprosum (ENL) has been considered as a classical example of an immune complex disease in man.1
In tuberculosis the relationship between tuberculin sensitivity, i.e., DTH mediated by T cells, and resistance has been a classical dispute 2-4 since Koch's5 original observations on tuberculin hypersensitivity.
In experimental murine leprosy M. lepraemurium multiplied during the first 4 weeks after foot pad inoculation. Later, no further increase in acid-fast bacilli (AFB) was observed in C57/BL mice. The bacilli became elongated and the morphological index decreased. These changes with arrest of bacterial multiplication were preceded by a local swelling of the foot pad due to the onset of an immune reaction associated with development of DTH6 In this model, development of protective immunity was thus closely associated with the development of DTH. In subsequent experiments, these two phenomena could be clearly dissociated,7 and repeated induction of a strong local DTH reaction at the site of inoculation did not influence the extent of the growth of bacilli.8
In studies of immune responses following infection with mycobacteria, an essential approach has been to identify individual immunogenic constituents of the mycobacteria followed by characterization of the type and strength of immune responses to individual antigenic components.
Antigenic structure of mycobacteria. The antigenic structure of M. leprae has been difficult to study for the obvious reason that the leprosy bacillus has not been reproducibly cultivated in vitro. The demonstration of extensive bacterial multiplication and systemic infection in the armadillo following inoculation with live M. leprae9 represented a major advance, since bacilli could now be isolated from the tissues of infected armadillos for studies of antigenic content. Subsequently, further major advances have been based on the use of recombinant DNA technology by which individual protein antigens of M. leprae can be produced in vitro and characterized with regard to reactivity with antibodies and T cells.10 These studies have been carried to considerable detail with characterization of distinct B-cell and T-cell reactive epitopes on individual molecules, e.g., on the 65-kDa heat-shock protein.11-14
It is most striking concerning the first monoclonal antibodies to M. leprae10 that at least 3 of the 5 reactive antigens were heat shock proteins. While heat-shock proteins show a striking conservation throughout evolution with great homology, e.g., between the 65-kDa heat-shock protein in procaryotes and the human homolog, they are strong immunogens during experimental immunization as well as upon infection.15 The high proportion of heat-shock protein reactive monoclonal antibodies in the early experiments is, however, also probably related to overrepresentation of heat-shock proteins in the initial M. leprae preparations used for immunization of the mice. Heat-shock proteins are cytoplasmic constituents, and thus are expected to be preserved in the M. leprae bacilli after purification from armadillo tissues; while recovery of other antigens would be lower since a great part of the bacilli in these tissues are dying or dead, resulting in considerable loss of some antigenic constituents. We would expect M. tuberculosis and M. bovis BCG grown on synthetic media to provide more constant supplies of antigen from closely related mycobacteria. This is evidently the case for secreted antigens and probably for cytoplasmic and membrane-bound antigens as well.
The complexity of the immunogenic constituents of mycobacteria is directly illustrated by crossed immunoelectrophoresis (CIE). Provided that a potent, polyvalent antibody preparation is available, concentrated culture fluids or bacterial sonicates produce patterns with a multitude of distinct precipitate lines corresponding to individual immunogenic components. Reference systems in CIE with numbering of individual components have been described for M. bovis BCG16 and for M. tuberculosis.17
Figure 1A shows a CIE pattern of M. bovis BCG culture fluid against polyvalent anti-BCG immunoglobulin in the top gel. Antigen 85, indicated with an arrow, gives a major precipitate with internal spurs. This precipitate is barely detectable in sonicates of washed bacilli. Figure 1B shows the pattern of the same BCG culture fluid against an antiserum raised by immunization with purified antigen 85. Three distinct precipitates are observed, labeled 85A, 85B, and 85C.
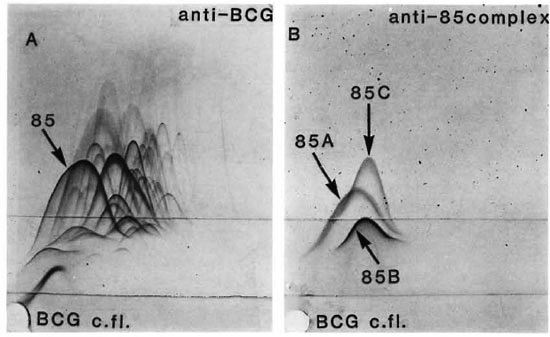
The figure. Crossed immunoelectrophoresis. Frame A-Antigen 85 complex gives a major precipitate inthe complex pattern produced with BCG culture fluid against polyvalent anti-BCG immunoglobulin in the topgel. Frame B-Pattern of the same BCG culture fluid with antiserum to the 85 antigen complex which consists of three components labeled 85A, 85B, and 85C.
Actively secreted and other antigens in culture fluids. Antigens may occur in culture fluids due to active secretion from the mycobacterial cells or because of release of cytoplasmic proteins into the culture medium after bacterial lysis. Conceptually, these two alternatives are clearly different. However, their distinction at the level of individual proteins has met with considerable difficulties.
Conditions are quite different concerning this matter in BCG and M. tuberculosis. In our experience, there is a greater tendency to bacterial lysis in cultures of BCG on Sauton medium than in cultures of M. tuberculosis. In the development of concepts and techniques, we found it essential to work with a particular M. tuberculosis culture fluid showing minimal bacterial lysis. In cultures of this kind, two major antigens showed entirely different quantitative patterns in comparative assays of culture fluids and sonicates of washed bacilli.18 Antigen 82, which corresponds to the 65-kDa heat-shock protein, was a dominating constituent of sonicates of washed bacilli, while it was barely detected in culture fluid. By contrast, antigen 85B was a major constituent of the culture fluid and barely detectable in sonicate of washed bacilli. For individual proteins this pattern may be expressed by a localization index (LI), which is defined as the following ratio:

This index can be determined directly only in cultures with minimal bacterial lysis. In cultures with significant bacterial lysis, the LI may be determined by means of correction for the degree of lysis by assay of marker cytoplasmic proteins that are released into the culture medium only by lysis during culture.19
Cytoplasmic proteins have an LI value close to 0, while secreted proteins were found to have markedly different values indicating differences in secretion efficiency. The highest LI value observed was for the MPB70 protein of M. bovis. The 85A, 85B, and 85C components of BCG had LI values of 150, 70, and 20, respectively, showing a distinctly different behavior for these three secreted proteins.
Antigen 85 complex. By CIE the antigen 85 complex was shown to consist of three distinct, although markedly crossreacting components, denoted 85A, 85B, and 85C.20 Subsequently, it was also demonstrated by CIE that the 85A, 85B, and 85C components were consistently present in all of seven substrains of BCG, including the culture fluid of cloned BCG Pasteur. The three proteins were isolated from a single culture fluid and the N-terminal sequence analysis of the purified proteins showed distinct differences, indicating that the components were encoded by separate genes.21 The sequence of the 85B component corresponded exactly to the deduced sequence of the "alpha-gene" cloned by Matsuo, et al.22 Borremans, el al.23 sequenced a gene encoding a 32-kDa protein of M. tuberculosis showing differences from the "alpha-gene" sequence. They interpreted these as species differences between BCG and M. tuberculosis. However, their sequence corresponded exactly to our sequence of BCG 85A. The differences were thus apparently due to differences in components rather than differences in species. We later tested the three components isolated from M. tuberculosis,24 finding their sequences to be identical to those of the 85A, 85B, and 85C components of BCG.
More recently, the gene encoding the 85C component of M. tuberculosis has been cloned and sequenced by Content, et al.25 The deduced amino-acid sequence is identical with the N-terminal sequence determined on the isolated protein. It was further demonstrated that the three genes coding for 85A, 85B, and 85C are located at different sites in the mycobacterial genome.
The proteins of the antigen 85 complex have been difficult to demonstrate directly in available M. leprae preparations, being sonicates of bacilli purified from armadillo tissue, and a major fraction of this group of antigens is expected to be released from the bacilli in vivo with further losses during the purification procedure. We have, however, been able to demonstrate crossreactive antigen in M. leprae sonicates by a sensitive double antibody ELISA designed for the demonstration of the 85B antigen of M. tuberculosis26 based on the use of the reference monoclonal antibody HYT27 which reacts with all three components of the M. tuberculosis antigen 85 complex.27-28
Recently, Thole, et al.29 have cloned a gene from M. leprae coding for a 327-amino-acid polypeptide, consisting of a consensus signal sequence of 38 amino acids followed by a mature protein of 289 amino acids most closely related to 85A. Southern blotting analysis indicated the presence of multiple genes of the 85 complex in the genome of M. leprae. The amino-acid sequence displays 75%-85% sequence identity with components of the antigen 85 complex from M. tuberculosis, M. bovis BCG, and M. kansasii.
A family of secreted proteins in mycobacteria. A series of other actively secreted proteins have recently been purified from M. tuberculosis culture fluid.24 One of them was termed MPT51 from its relative mobility in polyacrylamide gel electrophoresis (PAGE). This 27-kDa protein revealed a 60% homology with antigen 85B in the N-terminal amino-acid sequence, and polyclonal rabbit antibodies raised by immunization with highly purified MPT51 crossreacted extensively with the antigen 85 complex proteins in Western blotting. Crossreactivity was also observed with MPT64, another secreted protein30 which showed significant, but less homology. In addition, a striking internal homology was demonstrated between two different stretches within the 85B sequence and indicated between three stretches within the MPT64 molecule. Similar internal homologies have been essential in defining other protein families. Thus, a family of at least four secreted proteins with common structural features has been demonstrated in mycobacteria. MPT64 may also belong to this family.28
The proteins of the antigen 85 complex are fibronectin-binding proteins.31 The significance of this property for mycobacterial survival in vivo has not been established and, apart from fibronectin-binding, few data are available concerning the primary function of the 85 complex proteins. In Bacillus several secreted proteins are cleavage enzymes,32 and enzyme activities such as protease, mucinase, lipase and RNase have been detected in mycobacterial culture fluids.33
Significance of secreted antigens as immunogens after infection. The actively secreted proteins of M. tuberculosis are of great current interest and are intensively studied in several groups. An essential question is to what extent they are immunogenic and if immune responses to these proteins are of particular importance in relation to induction of protective immunity following infection or for development of clinical symptoms or complications during the ensuing disease.
Immunogenicity of antigen crossreactive with M. tuberculosis antigen 85B (denoted MPT59 in Nagai's original nomenclature) was initially demonstrated in armadillos and monkeys with systemic mycobacterial infection after inoculation with live M. leprae34 Antibodies to the 85-complex proteins have also been demonstrated in leprosy35-37 and tuberculosis38-40 in humans. Since these proteins are widely crossreactive,41 they would also be expected to be useful as a more "general" indicator of mycobacterial infection and have been employed successfully for this purpose in bovine tuberculosis and paratuberculosis.42 A sensitive ELISA based on an indicator system with protein G is particularly useful for the demonstration of antibodies in various species by a single assay since this protein binds strongly to immunoglobulin in more species than protein A.
Protective immunity in relation to specificity of immune responses. In infections in which protective immunity depends on humoral immunity, there is a close relationship between the antibody specificities and their significance for protection. This is widely recognized in different types of infection. Examples of this type of protective immunity are neutralization of bacterial exotoxins by toxin-neutralizing antibodies, facilitation of phagocytosis with destruction of microorganisms mediated by antibodies to capsular polysaccharides, and virus neutralizing antibodies preventing infectivity by reaction with antigenic determinants on the virus surface.
Immunity to infections with intracellular microorganisms depends upon the development of protective T-cell-mediated responses. In most infections of this kind the relationship between specificity and protective effect remains to be elucidated, and it has not been established whether particular antigens are essential for development of protective immunity in mycobacterial infections. In fact, according to current paradigms it is even difficult to understand how this could be the case. If antigen specificity is essential, this would probably imply that only certain antigens or groups of antigens tend to be presented on the surface of antigen-presenting cells (APC) in such a way that protective T-cell-mediated responses are elicited. Antigens actively secreted by mycobacteria during culture have been shown to be strong immunogens during infection inducing B-cell34-40 as well as T-cell responses. 40 43-45 We may expect this group of antigens to be of particular importance since similar secretion in vivo might favor release of this group of antigens into the extra-bacillary fluid in microcompartments within the infected cell, subsequently leading to presentation of peptides derived from these secreted proteins on the macrophage surface. It is not known to what extent this mechanism of antigen presentation dominates during the early stages after infection, the most essential time for the development of protective immunity. It is also possible that destruction of the cells containing the intracellular microorganisms by cytotoxic cells may result in the release of antigens followed by phagocytosis and processing by other cells in the immediate surroundings.
Additional studies, aiming at a better understanding of the mechanisms of antigen presentation after mycobacterial infection and to delineate the relationship between protective immunity and delayed-type hypersensitivity to mycobacterial antigens, remain truly essential matters for the improvement of our understanding of the development of either immunity or disease with the usual symptoms or complications such as reversal reactions with loss of nerve function in leprosy.
- Morten Harboe, M.D., Ph.D.
Professor of Medicine (Immunology)
University of Oslo
Institute of Immunology and Rheumatology
Fr. Qvamsgt. 1
0172 Oslo, Norway
1. Harboe, M. The immunology of leprosy. In: Leprosy. Hastings, R. C, ed. Edinburgh: Churchill Livingstone, 1985, pp. 53-87.
2. Youmans, G. P. Relationship between delayed hypersensitivity and immunity in tuberculosis. In: Tuberculosis. Philadelphia: W. B. Saunders Company, 1979, pp. 302-316.
3. Rook, G. A. W. and Stanford, J. L. The relevance to protection of three forms of delayed skin-test response evoked by M. leprae and other mycobacteria in mice. Correlation with classical work in the guineapig. Parasite Immunol. 1(1979)111-115.
4. Turk, J. L. Dissociation between allergy and immunity in mycobacterial infections. Lepr. Rev. 54(1983)1-8.
5. Koch, R. I. Weitere Mittheilungen über ein Heilmittel gegen Tuberculose. Dtsch. Med. Wochenschr. 46a(Suppl.)(1890)1029-1032.
6. Closs, O. Experimental murine leprosy: growth of Mycobacterium lepraemurium in C3H and C57/BL mice after foot pad inoculation. Infect. Immun. 12(1975)480-489.
7. Closs, O. and Løvik, M. Protective immunity and delayed-type hypersensitivity in C57BL mice after immunization with live Mycobacterium lepraemurium and sonicated bacilli. Infect. Immun. 29(1980)17-23.
8. Løvik, M. and Closs, O. Repeated delayed-type hypersensitivity reactions against Mycobacterium lepraemurium antigens at the infection site do not affect bacillary multiplication in C3H mice. Infect. Immun. 36(1982)768-774.
9. Kirchheimer, W. F. and Storrs, E. E. Attempts to establish the armadillo (Dasypus novemcinctus Linn.) as a model for the study of leprosy. I. Report of lepromatoid leprosy in an experimentally infected armadillo. Int. J. Lepr. 39(1971)693-702.
10. Young, R. A., Mehra, V., Sweetser, D., Buchanan, T., Clark-Curtiss, J., Davis, R. W. and Bloom, B. R. Genes for the major antigens of the leprosy parasite Mycobacterium leprae. Nature 316(1985)450-452.
11. Mehra, M., Sweetser, D. and Young, R. A. Efficient mapping of protein antigenic determinants. Proc. Natl. Acad. Sei. U.S.A. 83(1986)7013-7017.
12. Buchanan, T. M., Nomaguchi, H., Anderson, D. C, Young, R. A., Gillis, T. P., Britton, W. J., Ivanyi, J., Kolk, A. H. J., Closs, O., Bloom, B. R. and Mehra, V. Characterization of antibody-reactive epitopes on the 65-kilodalton protein of Mycobacterium leprae. Infect. Immun. 55(1987)1000-1055.
13. Thole, J. E. R., van Schooten, W. C. A., Keulen, W. J., Hermans, P. W. M., Janson, A. A. M., de Vries, R. R. P., Kolk, A. H. J. and van Emden, J. Use of recombinant antigens expressed in Escherichia coli K-12 to map B-cell and T-cell epitopes on the immunodominant 65-kilodalton protein of Mycobacterium bovis BCG. Infect. Immun. 56(1988)1633-1640.
14. Hajeer, A. H., Worthington, J., Morgan, K. and Bernstein, R. M. Monoclonal antibody epitopes of mycobacterial 65kDa heat-shock protein defined by epitope scanning. Clin. Exp. Immunol. (1992) (in press).
15. Harboe, M. and Quayle, A. J. Heat shock proteins: friend and foe? Clin. Exp. Immunol. 86(1991)2-5.
16. Closs, O., Harboe. M., Axelsen, N. H., Bunch-Christensen, K. and Magnusson, M. The antigens of Mycobacterium bovis. BCG, studied by crossed immunoelectrophoresis: a reference system. Scand. J. Immunol. 12(1980)249-263.
17. Wiker, H. G., Harboe, M., Bennedsen, J. and Closs, O. The antigens of Mycobacterium tuberculosis. H37Rv, studied by crossed immunoclectrophoresis. Comparison with a reference system for Mycobacterium bovis, BCG. Scand. J. Immunol. 27(1988)223-239.
18. Harboe, M., Wiker, H. G. and Nagai, S. Protein antigens of mycobacteria studied by quantitative immunologic techniques. Clin. Infect. Dis. 14(1992)313-319.
19. Wiker, H. G., Harboe, M. and Nagai, S. A localization index for distinction between extracellular and intracellular antigens of Mycobacterium tuberculosis. J. Gen. Microbiol. 137(1991)875-884.
20. Wiker, H. G., Harboe, M. and Lea. T. E. Purification and characterization of two protein antigens from the heterogenous BCG85 complex in Mycobacterium bovis BCG. Int. areh. Allergy Appl. Immunol. 81(1986)298-306.
21. Wiker, H. G., Sletten, K., Nagai, S. and Harboe, M. Evidence for three separate genes encoding the proteins of the mycobacterial antigen 85 complex. Infect. Immun. 58(1990)272-274.
22. Matsuo, K., Yamaguchi, R., Yamazaki, A., Tasaka, H. and Yamada. T. Cloning and expression of the M. bovis BCG gene for extracellular alpha antigen. J. Bacteriol. 170(1988)3847-3854.
23. Borremans, M., de Wit, L., Volckaert, G., Ooms, J., De Bruyn, J., Huygen, K., van Vooren, J.-P., Ste- landre, M., Verhofstadt, R. and Content, J. Cloning, sequence determination, and expression of a 32-kilodalton-protein gene of Mycobacterium tuberculosis. Infect. Immun. 57(1989)3123-3130.
24. Nagai, S., Wiker, H. G., Harboc, M. and Kinomo to, M. Isolation and partial characterization of main protein antigens in the culture fluid of Mycobacterium tuberculosis. Infect. Immun. 59(1991)372-382.
25. Content. J., de la Cuvellerie, A., de Wit, L., Vincent-Levy-Frébault, V., Ooms, J. and De Bruyn, J. The genes coding for the antigen 85 complexes of Mycobacterium tuberculosis and Mycobacterium bovis BCG are members of a gene family: cloning, sequence determination, and genomic organization of the gene coding for antigen 85-C of M. tuberculosis. Infect. Immun. 59(1991)3205-3212.
26. Wiker. H. G., Harboe, M., Nagai, S. and Bennedsen, J. Quantitative and qualitative studies on the major extra-cellular antigen of Mycobacterium tuberculosis H37Rv and Mycobacterium bovis BCG. Am. Rev. Respir. Dis. 141(1990)830-838.
27. Schou, C, Yuan, Z.-L., Andersen, B. and Bennedsen, J. Production and partial characterization of monoclonal hybridoma antibodies to Mycobacterium tuberculosis. Acta Pathol. Microbiol. Immunol. Scand. [C] 93(1985)265-272.
28. Wiker, H. G., Nagai, S., Harboe, M. and Ljungqvist, L. A family of crossreacting proteins secreted by Mycobacterium tuberculosis. Scand. J. Immunol. (1992) (in press).
29. Thole, J. E. R., Schöningh, R., Janson, A. A. M., Garbe, T., Corneliesse, Y. E., Clark-Curtis, J. E., Kolk, A. H. J., Ottenhoff, T. H. M., de Vries, R. R. P. and Abou-Zeid, C. Molecular and immunological analysis of a fibronectin-binding protein antigen secreted by Mycobacterium leprae. Mol. Microbiol. 6(1992)153-163.
30. Harboe, M., Nagai, S., Patarroyo, M. E., Torres, M. L., Ramirez, C. and Cruz, N. Properties of proteins MPB64, MPB70, and MPB80 of Mycobacterium bovis BCG. Infect. Immun. 52(1986)293-302.
31. Abou-Zeid, C, Ratlitf, T. L., Wiker, H. G., Harboe, M., Bennedsen, J. and Rook, G. A. W. Characterization of fibronectin-binding antigen released by Mycobacterium tuberculosis and Mycobacterium bovis BCG. Infect. Immun. 56(1988)3046-3051.
32. Sarvas, M. Protein secretion in bacilli. Curr. Top. Microbiol. Immunol. 125(1986)103-125.
33. Kannan, K. B., Katoch, V. M., Sharma, V. D. and Bharadwaj, V. D. Extracellular enzymes of mycobacteria. FEMS Microbiol. Lett. 48(1987)31-33.
34. Wiker, H. G., Harboe, M., Nagai, S., Patarroyo, M. E., Ramirez, C. and Cruz, N. MPB59, a widely cross-reacting protein of Mycobacterium bovis BCG. Int. areh. Allergy Appl. Immunol. 81(1986)307-314.
35. Rumschlag, H. S., Shinnick, T. M. and Cohen, M. L. Serological responses of patients with lepromatous and tuberculoid leprosy to 30-, 31-, and 32-kilodalton antigen of Mycobacterium tuberculosis. J. Clin. Microbiol. 26(1988)2200-2202.
36. Pessolani, M. C. V., Rumjanek, F. D., de Melo Marques, M. A., de Melo, F. S. F. and Samo, E. N. Serological response of patients with leprosy to a 28to 30-kilodalton protein doublet from early cultures of Mycobacterium bovis BCG. J. Clin. Microbiol. 27(1989)2184-2189.
37. Espitia, C, Sciutto, E., Bottasso, O., González-Amaro, R., Hernández-Pando, R. and Mancilla, R. High antibody levels to the mycobacterial fibronectinbinding antigen of 30-31kD in tuberculosis and lepromatous leprosy. Clin. Exp. Immunol. 87(1982)362-367.
38. Stroebel, A. B., Daniel, T. M., Lau, J. H. K., Leong, J.C.Y. and Richardson, H. Serologic diagnosis of bone and joint tuberculosis by an enzyme-linked immunosorbent assay. J. Infect. Dis. 146(1982)280-283.
39. van Vooren, J.-P., Drowart, A., de Cock, M., van Onckelen, A., D'Hoop, M. H., Yernault, J. C, Valcke, C. and Huygen, K. Humoral immune response of tuberculosis patients against the three components of the Mycobacterium bovis BCG 85 complex separated by isoelectric focusing. J. Clin. Microbiol. 29(1991)2348- 2350.
40. Wiker, H. G. and Harboc, M. The antigen 85 complex; a major secretion product of Mycobacterium tuberculosis. Microbiol. Rev. (submitted for publication).
41. Harboe, M., Mshana, R. N., Closs, O., Kronvall, G. and Axelsen, N. H. Cross-reactions between mycobacteria. II. Crossed immunoelectrophoretic analysis of soluble antigen of BCG and comparison with other mycobacteria. Scand. J. Immunol. 9(1979)115-124.
42. Harboe, M., Wiker, H. G., Duncan, J. R., Garcia, M. M., Dukes, T. W., Brooks, B. W., Turcotte, C. and Nagai, S. Protein G-based enzyme-linked immunosorbent assay for anti-MPB70 antibodies in bovine tuberculosis. J. Clin. Microbiol. 28(1990)913-921.
43. de Bruyn. J., Huygen, K., Bosmans, R., Fauville, M., Lippens, R., van Vooren, J.-P., Falmagne, P., Weckx, M., Wiker. H. G., Harboe, M. and Turneer, M. Purification, characterization and identification of a 32kDa protein antigen of Mycobacterium bovis BCG. Microb. Pathog. 2(1987)351-366.
44. Huygen, K.., van Vooren, J.-P., Turneer, M., Bosmans, R., Dierckx, P. and de Bruyn, J. Specific lymphoproliferation. gamma interferon production, and serum immunoglobulin G directed against a purified 32 kDa mycobacterial protein antigen (P32) in patients with active tuberculosis. Scand. J. Immunol. 27(1988)187-194.
45. Andersen, P., Askgaard, D., Ljungqvist, L., Bentzon, M. W. and Heron, I. T-Cell proliferative response to antigens secreted by Mycobacterium tuberculosis. Infect. Immun. 59(1991)1558-1563.