- Volume 59 , Number 2
- Page: 262–70
Determination of in vivo and in vitro drug effects of mycobacteria f rom the mass spectrometric analysis of single organisms
ABSTRACT
Laser microprobe mass analysis of single bacterial organisms allows the determination of their intrabacterial ratio of sodium-to-potassium ions and the registration of fragment ions originating f rom their organic bacterial cell matrices as mass fingerprint spectra. It has been established previously that the intrabactcrial cation ratio provides information on the physiological state of an individual bacterial cell. In the present experiments it is also shown, with different cultivable mycobacterial species and strains (drug sensitive and resistant) exposed to various drugs, that data derived f rom the evaluation of the mass fingerprint spectra reflect changes in the degree of impairment. The analysis of Mycobacterium leprae derived f rom a limited number of skin biopsies of lepromatous/borderlinc lepromatous leprosy patients under World Health Organization-recommended multiple-drug therapy (WHO/MDT) showed impairment of the organisms with both of the methods of measurement in proportion to the duration of treatment except in one case. In one M. leprae population f rom a patient who had been treated for 19 months, the fingerprint evaluation gave the first evidence for an insufficient response to treatment. This was further confirmed by the unusual frequency distribution of the Na+,K+ ratios which revealed the existence of two subpopulations, one impaired and one unimpaired.RÉSUMÉ
L'analyse de masse par microsonde laser d'organismes bactériens isolés permet la détermination du ratio intrabactérien des ions sodium/potassium, et l'enregistrement de fragments ioniques originaires de leurs matrices cellulaires organiques en tant qu' empreintes digitales du mélange obtenues par spectrométrie de masse. Il a été établi auparavant que le ratio des cations intrabactériens fournit des informations sur l'état physiologique d'une cellule bactérienne individuelle. L'expérimentation présente montre également en utilisant difiêrcntcs espèces et souches (sensibles et résistantes aux médicaments) de mycobactérics cultivables, que les données obtenues par l'évaluation des empreintes digitales obtenues par spectrométrie de masse reflètent difiérents degrés d'altération. L'analyse de Mycobacterium leprae obtenus à partir d'un nombre limité de biopsies cutanées de patients présentant une lèpre lé-promateusc/borderlinc lépromatcusc soumis à une polychimiothérapie telle que recommandé par l'OMS (PCT/OMS) a montré une altération des organismes avec les deux méthodes de mesure, et ceci proportionnellement à la durée de traitement, à l'exception d'un cas. Dans une population de M. Ieprae d'un patient qui avait été traité pour 19 mois, l'évaluation par empreintes fut la première à mettre en évidence une réponse insuffisante au traitement. Ceci fut confirmé par la suite par la distribution de fréquences inhabituelle des ratios NA+ et K+, qui révéla l'existence de deux sous-populations, l'une altérée, et l'autre non.RESUMEN
El microanálisis de masas con rayos laser de organismos bacterianos individuales, permite la determinación de su relación sodio/potasio y el registro del espectro de los fragmentos iónicos que se originan de la matriz orgánica celular. Ya antes se estableció que la relación de cationes proporciona información sobre el estado fisiológico de una célula bacteriana individual. En el presente estudio se muestra que el micro-análisis de masas con rayos laser revela también el grado de alteración inducido por varias drogas en diferentes micobacterias cultivables, sensibles y resistentes a los drogas. El análisis de las células de Mycobacterium leprae aisladas de un número limitado de biopsias de piel de pacientes LL/BL sujetos al tratamiento con múltiples drogas recomendado por la Organización Mundial de la Salud (OMS), demostró la alteración de los microorganismos, por los dos métodos de medición, en proporción a la duración del tratamiento, excepto en un caso. El microanálisis de masas dio la primer evidencia de una insuficiente respuesta al tratamiento en un paciente que había sido tratado durante 19 meses. Esto fue posteriormente confirmado por la anormal distribución de la relación Na+, K+, que reveló la existencia de 2 subpoblacioncs, una dañada y otra no alterada.The standard microbiological methods for the determination of drug effects on bacteria in vivo or in vitro are usually based on data obtained as averages of a large number of organisms and/or on the ability of the organisms to multiply. Mycobacterium leprae does not fulfill these requirements. In spite of tremendous efforts of several laboratories over many decades, cultivation in artificial broth media has not been achieved (3). Thus, the usual procedures for monitoring therapy and drug development employ animal models (2,8,11), and these have some inherent restrictions; firstly, because results are available only after long periods of time (typically 10 months) and, secondly, one has to rely on the assumption that the metabolisms of the animal and human hosts are comparable. Therefore, several attempts have been made to develop alternative methods which would a) avoid animal experiments, b) require small numbers of bacteria, and c) most importantly, provide the results in considerably shorter time (1).
Our approach is based on the mass spectrometric analysis of a limited number of single organisms (typically between 100 and 300). These are isolated from infected tissue or - in case of cultivable bacteria - from broth media for the determination of their intracellular ratio of sodium and potassium ions (Na+,K+-ratio) and of their so-called mass fingerprints comprising fragment ions of the organic matrix of the bacteria. The intracellular cation ratio has been shown to be a sensitive indicator of the physiological state of bacteria (9). The mass fingerprints reflect differences among bacterial samples due to either environmental influences or species-specific characteristics (10).
In a first extended study with 34 leprosy patients treated with 4,4'-diaminodiphenyl sulfone (DDS), we have established that the drug influences the intracellular Na+,K+-ra-tios as well as the mass fingerprints of M. leprae in vivo. The fingerprint evaluation provided a separation into DDS-treated and -untreated samples (14). The sample specificities, however, leading to this discrimination are presently not known. In principle, they can be related to: a) the mere presence of drug molecules; b) drug-induced changes of the bacterial cell matrix, which might even be drug- or species-specific; or c) drug-induced impairments, which could possibly correlate with changes in the intra-bacterial Na+,K+-ratio.
We report on experiments performed to establish the possible parameters responsible for the discrimination between various bacterial samples on the basis of their mass fingerprint spectra. For this, we have studied, with cultivable mycobacteria, the similarity relationships between different samples of one species exposed for various times to one drug and between samples of different species, each treated with three different drugs, respectively. These experiments revealed, in correlation with the results from the measurement of the cation ratios, that the degree of impairment is the dominant parameter for the fingerprint differentiation. These findings allow the recognition of bacterial drug resistance on the basis of similarity relationships between fingerprint spectra. This is demonstrated in an experiment in which isonicotinic acid hydrazide (INH)-sensitive and -resistant strains were exposed to various concentrations of INH. We further report on the application of these methods to M. leprae isolated from patients under the multidrug therapy proposed by the World Health Organization (WHO/ MDT), demonstrating that the single cell mass analysis of M. leprae can provide fast experimental proof of suspected drug resistance.
MATERIALS AND METHODS
Cultivable bacteria
Strains. M. smegmatis no. 1546/85, M. avium no. 7901/87, and M. vaccae SN 912 were used (Division of Microbiology, Borstel Research Institute, Borstel, Germany).
Growth. Stock cultures were maintained on Lowenstein-Jensen slants (DIFCO Laboratories, Detroit, Michigan, U.S.A.)at room temperature; at 2-month intervals fresh slants were inoculated and incubated at 37ºC until visible growth.
Preliminary culture: 100 ml of modified Dubos broth (16)-containing 0.35% w/v bovine serum albumin fraction V and 6.25 g/1 disodium phosphate-were inoculated with 106 ml-1 and allowed to grow up to the early stationary phase (107- 108 ml-1) at 37ºC.
For the main cultures, modified Dubos broth was inoculated from preliminary cultures with 5 x 105 ml-1 and incubated at 37ºC in Erlenmeyer flasks on a shaking table. Growth was controlled by counting the total number of bacteria (Neubauer counting chamber) every 2 to 4 hr, depending on the generation time.
The INH-resistant M. vaccae was produced by sequential passage of M. vaccae (sensitive strain) in the presence of increasing concentrations of INH until the organisms were able to grow in the presence of 150 μg/ml-1 INH. The suspension of the resistant bacteria was plated on Dubos oleic albumin agar (15) containing 100 μg ml-1 INH to obtain single colonies of resistant bacteria. One of these colonies was isolated and multiplied under the same conditions as the stock culture for the INH-resistant strain of M. vaccae.
Treatment. The following drugs were used: rifampin (RMP), trimethoprim (TMP), polymyxin B sulfate (PMB), and isonicotinic acid hydrazide (INH) (Sigma Chemicals, Deisenhofen, Germany). The drugs were added in the exponential growth phase at about 5 x 106 ml-1 from stock solutions. Solvent concentrations (DMSO for TMP, distilled water/HCl for RMP, distilled water for PMB and INH) did not exceed the final concentration of 0.1%. Drug concentrations were referred to 50% inhibitory concentration (I50) values as determined by growth inhibition experiments with M. smegmatis or M. vaccae (INH-sen-sitive strain) under the same growth conditions as before. The I50 values determined this way with M. smegmatis were 5.5 μg ml-1 for TMP, 4.5 μg ml-1 for PMB, 1.3 μg ml-1 for RMP, and 3 μg ml-1 for INH. The 150 for INH determined with M. vaccae (INH-sensitive strain) was identical with that determined for M. smegmatis. After drug addition, the cultures were incubated until the untreated control had grown to the early stationary phase. Samples were taken for single cell mass analysis and for total and variable cell counting at times indicated in the respective figures.
M. leprae
Patients and treatment. Skin biopsies were collected from multibacillary leprosy patients (LL/BL cases) at the All Africa Leprosy and Rehabilitation Center (ALERT) in Addis Ababa, Ethiopia, before and at different times after the beginning of treatment and shipped on dry ice to the Forschungsinstitut Borstel, where M. leprae were isolated and prepared for single cell mass analysis. This study covered four inpatients treated according to the WHO/MDT, including DDS (100 mg once a day), RMP (600 mg once a month), and clofazimine (50 mg once a day, 300 mg once a month). The patients were under clinical control throughout the study.
Isolation and preparation procedures.
The preparation procedure for M. leprae followed a modification of the protocol of Dhople (4). Briefly, it includes the following steps: removal of epidermis; homogeniza-tion in phosphate buffered saline (PBS); removal of intact tissue via centrifugation; decontamination of M. leprae from remaining tissue by treatment with proteolytic enzymes (collagenase, chymotrypsin, trypsin); washing in PBS; decontamination from other bacterial genera with N-acetyl-L-cystcine and NaOH (7); removal of extracellular sodium and potassium by two washing and centrifugation steps in distilled water; transfer of one drop of the concentrated bacterial suspension to a Formvar-coated copper mesh (as used in electron microscopy); and draining of excess fluid with tissue paper to achieve a widespread distribution of bacteria, which allows the laser evaporation of one single cell at a time.
Laser microprobe mass analysis (LAM-MA). This technique has been described in detail elsewhere (6). Briefly, in the LAM-MA instrument a high-energy, ultraviolet laser pulse is focused through the objective of a light microscope onto the sample to be analyzed and, in our experiment, evaporates one bacterial organism at one shot. The positive atomic and molecular (fragment) ions produced are registered by means of a time-of-flight mass spectrometer. The detection limits of the instrument for sodium and potasssium are lower than 10-19 g out of an analyzed mass of 10-12 g (0.1 ppm).
Data evaluation. The intracellular Na+,K+-ratio was determined from the intensities of the 23Na+- and 39K+-mass peaks from 300 single-cell measurements. The measured ratio does not necessarily reveal the true intracellular ratio because of different ionization efficiencies for the two cations. from these, relative cumulative distributions (RCDs) were calculated, permitting the determination of the percentage of bacteria having a Na+,K+-ratio below a given limit.
The mass spectra of the organic fragment ions comprise a large number of peaks in the mass range m/z 60 to 250. The patterns of the mass peaks are characteristic for the bacteria of a given sample and can thus be taken as mass fingerprints. To overcome unavoidable variations from shot to shot of individual organisms, 120 spectra from each sample are registered and summarized in four averaged spectra over 30 single-cell spectra each. Because it cannot be judged by direct visualization whether the differences between patterns of spectra of different samples are significantly larger than the differences between those of the same sample, computerized statistical methods (10) are used to calculate numerical similarity relationships between the various averaged spectra. The results are plotted in a two-dimensional graph representing the numerical relationships between the various spectra. In these maps each averaged spectrum is represented by a point, and points belonging to the same sample are interconnected. A high degree of similarity between two spectra is expressed by a short distance between the respective points. The underlying mathematical algorithm of this procedure is called multidimensional scaling (13).
RESULTS
Cultivable bacteria. In respect to our intention to find out which information can be derived from the evaluation of mass fingerprint spectra, in our first experiment we have treated M. smegrnatis with 5 x I50 TMP for different times. Bacterial growth of the control population and of the treated culture is shown in Figure 1A. At different times after drug addition, samples were taken and prepared for single cell mass analysis to determine the intracellular cation ratios and to register the fingerprint patterns.
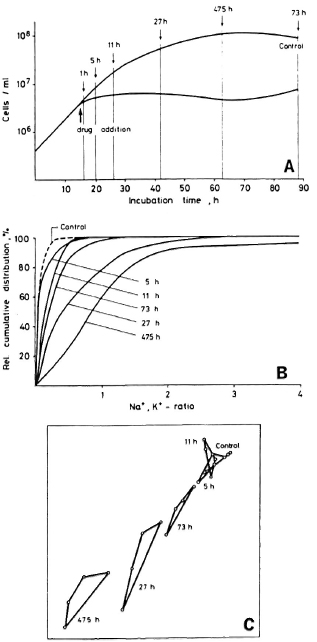
Fig. 1. Growth kinetics (A) and relative cumulative distributions of the intracellular Na+,K+-ratios (B) of an untreated culture of M. smegmatis and of cultures treated with 5 x I50 of TMP for various times. Similarity relationships between fingerprint spectra of the same samples are shown in a two-dimensional nonlinear map (C).
Relative cumulative distributions of the Na+,K+-ratios obtained from 300 single cells for each sample taken from the controls and the drug-treated culture are depicted in Figure 1B. Only one control at one time is included because changes in the distributions during the complete experiments were only minimal. Under the influence of TMP, the shape of the distributions is changed and the median (50%-value) is shifted to higher values of the cation ratio up to an incubation time of 47.5 hr and the reversal of this effect at 73 hr is observed.
The evaluation of the fingerprint spectra in a two-dimensional nonlinear map of the similarity relationships between the untreated control and the samples treated with TMP for different times is given in Figure 1C. The four averaged spectra belonging to the same sample are interconnected. The distances between the areas represent the numerical similarities between the different samples. from this representation it can be taken that, for instance, the control and the sample treated with TMP for 47.5 hr are highly dissimilar; whereas the control and those samples treated with TMP for 5 hr and 11 hr, respectively, are closely related.
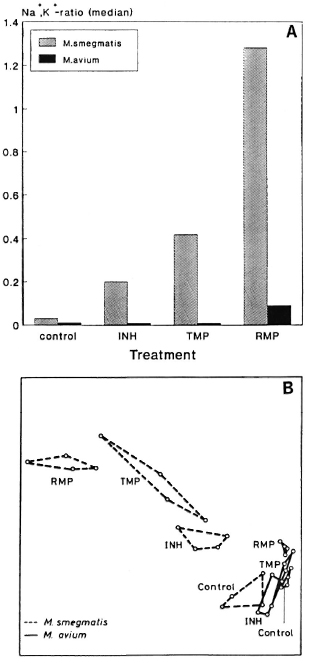
Fig. 2. Medians of distributions of intracellular Na+,K+-ratios of M. smegmatis and M. avium cultures treated with 2 x I50 of TMP, RMP and INH, respectively, for 10 generation times (A), and the two-dimensional nonlinear map of the similarity relationships between the same samples (B).
These findings back the respective data from the cation measurements (Fig. 1B).
To elucidate whether and to what extent species-specific influences are superimposed by drug-specific influences, we have compared the influences of three different drugs (TMP, RMP, INH) on two different mycobacterial species (the fast-growing M. smegmatis and the slow-growing M. avium). Cultures of these species were incubated for 10 generation times with 2 x I50 TMP, RMP or INH, respectively (as determined by growth inhibition for M. smegmatis).
In Figure 2A the medians of the distributions of the cation ratios obtained from 300 single cells in each case are plotted for this experiment, showing that the fast-growing M. smegmatis is more sensitive to each one of the drugs than the slow-growing M. avium and that RMP is the most effective drug against both species. Figure 2B shows the two-dimensional nonlinear map of the similarity relationships between the controls of the two species and the differently treated cultures. Both control cultures of M. avium and M. smegmatis as well as those samples of M. avium treated with the different drugs form one single cluster; the INH-, TMP- and RMP-treated M. smegmatis cultures are well separated from each other and from the corresponding control.
In an experiment with an INH-sensitive and an INH-resistant strain of M. vaccae, the influence of two different concentrations of INH (20 x I50 and 30 x l50, both values referred to the sensitive strain) on the two measuring parameters was investigated in order to test their applicability to the recognition of drug resistance. The results are summarized in Figure 3, showing significant concentration-dependent changes in the Na+,K+-ratios (A) as well as in the similarity relationships between the mass fingerprints (B) only for the INH-sensitive strain.
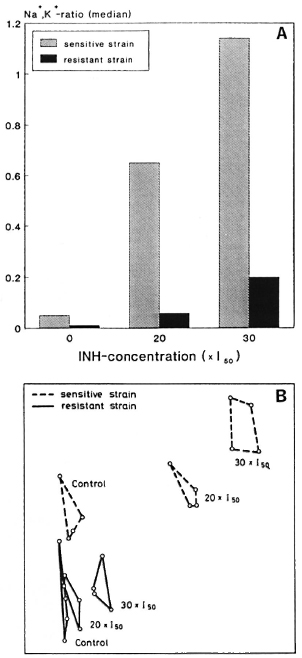
Fig. 3. Medians of distributions of intrabactcrial Na+, K+-ratios of an INH-sensitive and an INH-resis-tant strain of M. vaccae not treated and treated with 20 x l50 and 30 x I50 INH, respectively, (A) and two-dimensional nonlinear map of the similarity relationships between fingerprint spectra of the same samples (B).
M. leprae. To find out whether similarly detailed information as derived from the in vitro experiments with cultivable mycobacterial species can also be obtained for in vivo drug therapy of leprosy, we applied the described method to M. leprae isolated from biopsies of a limited number of patients treated with WHO/MDT for different durations.
from Figure 4A it is obvious that the medians of the Na+,K+-ratios of the isolates from patients before and at different times during treatment show an increase according to the duration of treatment. The two-dimensional nonlinear map of the numerical similarity relationships between these M. leprae populations reveals clustering of the untreated versus the treated samples (Fig. 4B). The cluster formed by the samples obtained from treated patients is arranged in a way roughly corresponding to the duration of treatment, except for that isolated from a patient having been under therapy for 19 months. In this case, the frequency distribution of the intracellular Na+,K+-ratios obtained from 500 bacteria (Fig. 5) clearly shows that the bacterial population consists of two subpopulations.
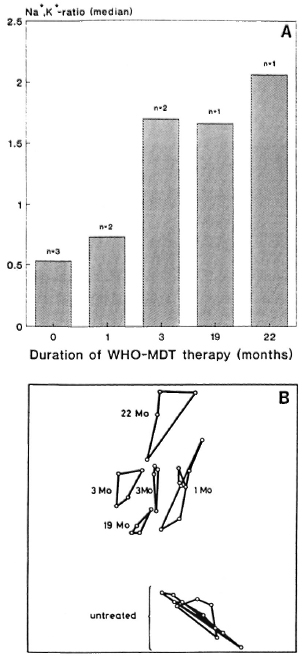
Fig. 4. Medians of distributions of intrabactcrial Na+,K+-ratios of M leprae isolated from human skin biopsies of patients (n = number of patients) before and at various times after the onset of treatment with WHO/MDT (A) and two-dimensional nonlinear map of the similarity relationships between the fingerprint spectra of M. leprae isolated from the same patients (B).
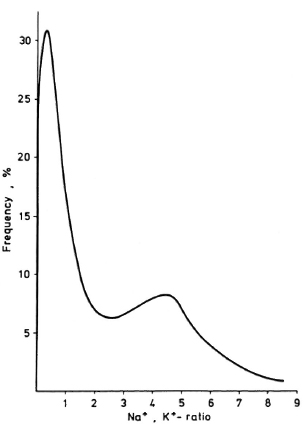
Fig. 5. Frequency distribution of intracellular Na+,K+-ratios of M. leprae isolated from a patient's biopsy after 19 months of treatment with WHO/MDT.
DISCUSSION
Mass spectra of single bacterial cells comprise ion signals from intrabactcrial cations as well as from fragment ions of the organic cell matrix. It could be shown that, from the former, direct information on the physiological state of single organisms can be derived, in particular from the ratio of the sodium and potassium concentrations (9). The measurement of this parameter allows one to monitor drug-induced impairments of bacterial populations via the analysis of some hundred single organisms per sample. Upon the administration of a drug to a bacterial culture, the patterns of mass fingerprints obtained from the bacterial organisms also change, as can be visualized by applying multivariate pattern recognition techniques (10). The experiments performed with cultivable mycobacteria now clearly demonstrate that these changes correlate with changes in the Na+,K+-ratio and, with that, reflect mainly the degree of impairment. This becomes obvious from the experiment with M. smegmatis treated with 5 x I50 TMP for various times: In the two-dimensional nonlinear map of the similarity relationships (Fig. 1C), the distance between the various samples and the control-each represented by a tetragon -increases with the duration of treatment up to 47.5 hr. The 73-hr sample, however, shows a higher degree of similarity with the untreated control, indicating a recovery of the bacteria. A corresponding effect is observed from the medians of the distributions of the Na+,K+-ratios (Fig. IB). Both parameters, the cation ratio and the fingerprints, show this recovery more pronouncedly than the total cell number (Fig. 1A).
The correlation between the shifts in the similarity relationship and in the medians of the distributions of the cation ratios is also confirmed in an experiment in which two different mycobacterial species were treated with three different drugs (Fig. 2). Thus, the dominant criterion for a separation of the various samples by multivariate analysis of their fingerprint spectra is the degree of impairment and not species- or drug-specific characteristics. In particular, the separation between the samples does not depend on the amount of the administered drug; otherwise, the fingerprints of M. smegmatis and M. avium should show comparable similarity shifts related to the control. Therefore, it should be possible to recognize bacterial drug resistance via the evaluation of mass fingerprints. Thus, the treatment of an INH-sensitive and INH-resistant strain of M. vaccae with two different concentrations of INH (Fig. 3) leads, in the case of the sensitive strain only, to marked changes in the similarity relationships of the treated samples relative to the control. This fact is also reflected in the medians of the Na+,K+-ratios.
from the above-described experimental results it may be concluded that the evaluation of mass fingerprint spectra offers an independent method for monitoring drug-induced bacterial impairment.
On the basis of these findings the results obtained from the fingerprint evaluation of M. leprae isolated from patients treated with WHO/MDT allow one to draw conclusions on the effectiveness of a therapy from the positions of the samples in the two-dimensional map (Fig. 4B). However, for this one must consider that the results from in vivo experiments may be influenced by the crude isolation procedure and by the variations in the drug response between different patients. In Figure 4B, the samples are arranged roughly according to the duration of treatment except for the sample obtained from a patient after 19 months of treatment. According to the findings from the in vitro experiments discussed above, this would imply an insufficient response of M. leprae to the WHO/MDT because drug uptake was supervised. The suspicion of WHO/MDT resistance was further backed by clinical observations. The patient who was put on WHO/MDT had developed new nodules under DDS monotherapy. However, also under the multidrug regimen the bacterial index (BI) started to increase after 1 year of treatment and new nodules appeared. The median of the Na+,K+-ratios of bacteria isolated from this patient fits well into the time dependent increase of this parameter as determined from the bacterial isolates from the other patients (Fig. 4A). The frequency distribution of the Na+,K+-ratios (Fig. 5), however, clearly reveals the existence of two subpopulations, an impaired and an unimpaired, the latter having a median typical for an untreated control. The median of the distribution of the cation ratios determined for the entire population does not reflect this situation. This example demonstrates that integral methods can supply reliable information only in the case of homogeneous bacterial populations. This implies that drug resistance cannot be detected with those methods when the drug resistance is in an early state of development.
A particular problem still unsolved in connection with the control of therapy is the determination of the percentage of viables in a human skin biopsy. This value cannot be derived from the measurements described above, because they allow only statements on changes in the physiological state relative to the untreated control. The quantification of the percentage of viables requires the determination of the intracellular Na+,K+-ratio up to which M. leprae are viable (limiting value).
We approached this problem in the following way: Under the presumption that the data obtained from experiments with cultivable mycobacteria can be transferred to M. leprae, the upper limit of the Na+,K+-ratios of viable organisms can be estimated by correlating, for the same experiment, the relative cumulative distributions of the Na+,K+-ratios with the percentage of viables determined as colony forming units per total cell number. So far, the upper limit of viability has been determined in a number of experiments with M. smegmatis treated with TMP and PMB (data not shown), resulting in the average value of 0.47 ± 0.03 (mean ± S.E.M., N = 25). from the evaluation of the relative cumulative distributions obtained from the biopsies of 31 untreated patients, this value would imply that on the average 30%-40% of untreated M. leprae arc viable. This value seems to be reasonable considering the marked changes in the measuring parameters during treatment which become visable from the analysis of only a few hundred single organisms. Also, in other alternative techniques, such as fluorescence and ATP assays, large changes in the respective measuring parameters upon drug interaction are observed which could not be understood if the percentage of viables were only in the order of a few percent as derived from the mouse foot pad test (5,16) and the morphological index (8,12). Nevertheless, the limiting value of 0.47 has to be considered preliminary, and should be proven in further experiments and other bacterial species and drugs under different growth conditions.
Acknowledgment. The authors acknowledge the support by the AHRI/ALERT Research Committee (Addis Ababa, Ethiopia). They are indebted to Mrs. G. Nagorny for her skillful assistance. This work was supported by the Federal Minister for Research and Technology (grant no. 03 8667/4) and by the German Leprosy Relief Association.
REFERENCES
1. Barclay, R. and Wheeler, P. R. Metabolism of mycobacteria in tissue. In: The Biology ofthe Mycobacteria. Vol. 3. Clinical Aspects of Mycobacterial Disease. Ratledgc, C, Stanford, J. and Grange, J. M., eds. London: Academic Press, 1989, pp. 37-106.
2. Colston, M. J. and Levy, L. Infection of other experimental animals with Mycobacterium leprae. Int. J. Lepr. 55Suppl.(1987)896-898.
3. Dhople, A. M. The status of in vitro cultivation of Mycobacterium leprae. Med. Sci. Res. 15(1987)599-603.
4. Dhople, A. M. and Storrs, E. E. Adcnosinetri-phosphate content of M. leprae: effect of purification procedures. Int. J. Lepr. 50(1982)83-89.
5. Grosset, J. H. Recent developments in the field of multidrug therapy and future research-in chemotherapy of leprosy. Lepr. Rev. 57(1986)223-234.
6. Heinen, H. J., Hillenkamp, F., Kaufmann, R., Schroeder, W. and Wechsung, R. LAM MA: a new laser microprobe mass analyser for biomed-icinc and material analysis. In: Recent Developments in Mass Spectrometry in Biochemistry and Medicine. Frigerio, A. and McCamish, M., eds. Amsterdam: Elsevier Science Publ., 1980, vol. 6, pp. 435-459.
7. K.uhica, G. P., Dye, W. E., Cohn, M. L. and Mid-dlebrook, G. Sputum digestion and decontamination with N-acetyl-L-cystein-sodium hydroxide for culture of mycobacteria. Am. Rev. Respir. Dis. 87(1963)775-779.
8. Levy, L. Application of the mouse foot-pad technique in immunologically normal mice in support of clinical drug trials, and a review of earlier clinical drug trials in lepromatous leprosy. Int. J. Lepr. 55Suppl.(1987)823-829.
9. Lindner, B. and Seydel, U. Mass spectrometric analysis of drug-induced changes in Na' and K.' contents of single bacterial cells. J. Gen. Microbiol. 129(1983)51-55.
10. Lindner, B. and Seydel, U. Results on taxonomy and physiological state of bacteria derived from laser-induced single cell mass analysis. J. Phys. Colloq. (France) 45C-2(1984)565-568.
11. McDermott-Lancaster, R. D., Ito, T., Kohsaka, K.., Guelpa-Lauras, C.-C. and Grosset, J. H. Multiplication of Mycobacterium leprae in the nude mouse, and some applications of nude mice to experimental leprosy. Int. J. Lepr. 55Suppl.(1987)889-895.
12. Padma, M. N. Morphological index in untreated cases of leprosy. Lepr. India 48(1976)787-789.
13. Seydel, U., Haas, M. and Lindner, B. In vitro drug effects monitored by single bacterial cell mass analysis with LAMMA. In: Microbeam Analysis - 1988. Newbury, E., ed. San Francisco: San Francisco Press, 1988, pp. 353-355.
14. Seydel, U., Lindner, B. and Dhople, A. M. Results from cation and mass-fingerprint analysis of single cells and from ATP measurements of M. leprae for drug sensitivity testing: a comparison. Int. J. Lepr. 53(1985)365-372.
15. Vera, H. D. and Dunoff, M. Culture media. In: Manual of Clinical Microbiology. 2nd ed. Lcn-nette, E. H., Spaulding, E. H. and Truant, J. P., eds. Washington: American Society for Microbiology, 1974, p. 901.
16. Welch, T. M., Gelber, R. H., Murray, L. P., Herman, N. G., O'Neill, S. M. and Levy, L. Viability of M. leprae after multiplication in mice. Infect. Immun. 30(1980)325-328.
1. M.Sc; Forschungsinstitut Borstel, Division of Biophysics, Parkallee 10, D-2061 Borstel, Germany.
2. Ph.D.; Forschungsinstitut Borstel, Division of Biophysics, Parkallee 10, D-2061 Borstel, Germany.
3. Ph.D., Professor, Forschungsinstitut Borstel, Division of Biophysics, Parkallee 10, D-2061 Borstel, Germany.
4. M.D., Ph.D.; All-Africa Leprosy and Rehabilitation Center (ALERT), P.O. Box 165, Addis Ababa, Ethiopia.
5. M.D., D.D.V., Ph.D., All-Africa Leprosy and Rehabilitation Center (ALERT), P.O. Box 165, Addis Ababa, Ethiopia.
Reprint requests to Prof. Seydel.
Received for publication on 26 July 1990.
Accepted for publication in revised form on 27 November 1990.