- Volume 58 , Number 4
- Page: 681–9
The disease of CBA and BALB/c mice that follows inoculation of a small number of Mycobacterium lepraemurium into the hind foot pad
ABSTRACT
To learn if the lack of an immune response in mice infected with Mycobacterium lepraemurium (MLM) was a consequence of the organisms, we studied the disease that followed inoculation of < 5000 organisms into the hind foot pads of CBA and BALB/c mice. The mice of both strains demonstrated a rapid increase of bacterial numbers soon after inoculation, with a slowing of the rate of multiplication once the number of organisms per foot pad passed 3 x 107. By 1 year after inoculation, the numbers of organisms had reached levels > 1011 in the spleen and liver, and > 108 in the femoral bone marrow. In mice that had been inoculated with as few as 5 MLM or 50 MLM, the organisms had multiplied to numbers > 108 in the foot pads and to > 109 in the spleens, suggesting that the ID50 of viable MLM may be < 5 organisms per foot pad. No protection against superinfection could be demonstrated. On the other hand, initial multiplication of MLM in the foot pads was followed virtually immedi-ately by the death of at least 97% of the organisms.RÉSUMÉ
Afin d'étudier si le manque de réponse immunitaire chez la souris infectée par Mycobocterium lepraemurium (MLM) était causée par les organismes, nous avons étudié la maladie qui suivait l'inoculation d'une quantité inférieure ou égale à 5000 organismes dans le coussinet plantaire postérieur de souris CBA et BALB/c. Les souris des deux espèces ont montré une augmentation rapide du nombre de bactéries précocement après l'inoculation, avec un ralentissement du taux de multiplication lorsque le nombre d'organismes par coussinet plantaire était passé à 3 x 107. Un an après l'inoculation, le nombre d'organismes avait atteint des niveaux supérieurs ou égaux à 1011 dans la rate et le foie, et à 108 dans la moelle osseuse fémorale. Chez des souris qui avaient été inoculées avec des quantités aussi faibles que 5 à 50 MLM, les organismes s'étaient multipliés jusqu'à des quantités supérieures à 108 dans les coussinets plantaires, et supérieures ou égales à 108 dans la rate, suggérant que la LD50 de MLM viables peut être inférieure ou égale à 5 MLM par coussinet plantaire. Aucune protection n'a pu être démontrée vis-à-vis de la surinfection. D'autre part, la multiplication initiale de MLM dans les coussinets plantaires fut suivie pratiquement immédiatement de la mort d'au moins 97% des organismes.RESUMEN
Para saber si la falta de respuesta inmune en ratones infectados con Mycobocterium lepraemurium (MLM) es una consecuencia de la enfermedad, se inocularon <5000 organismos en las almohadillas de las patas posteriores de ratones CBA y BALB/c. Los ratones de ambas cepas mostraron un rápido incremento en el número de bacterias poco tiempo después de la inoculación, con una disminución en la velocidad de multiplicación una vez que el número de organismos por almohadilla plantar pasó de 3 x 107. Un año después de la inoculación, los números de organismos alcanzaron niveles > 1011 en el bazo y en el hígado y > 108 en la médula ósea femoral. En los ratones inoculados con 5 ó 50 MLM, los organismos se multiplicaron a números mayores de 108 en las almohadillas plantares y a números > 108 en los bazos, sugiriendo que la LD50 de MLM viables puede ser de < 5 organismos por almohadilla plantar. No se pudo demostrar protección contra la superinfección por MLM. Por otro lado, la multiplicación inicial del MLM en la almohadilla plantar fue seguida casi inmediatamente por la muerte de cuando menos el 97% de los organismos.Systemic infection by Mycobacterium lepraemurium (MLM) of mice of several inbred strains results in a chronic and, finally, lethal disease process, characterized by multiplication of the organisms to very large numbers and massive accumulation of MLM-laden macrophages in the spleen, liver and bone marrow (2), and is accompanied by the loss of T-lymphocyte-mediated immune responses to the organism (15). The severity of the disease following subcutaneous injection of the organisms has been reported to vary widely among mice of different strains (3). In C3H mice of certain substrains, for example, subcutaneous infection leads to systemic, progressive disease; whereas the infection of C57BL mice is self-limited (5).
Our interest in the process that results from subcutaneous inoculation of mice with small numbers of MLM was stimulated by an unexpected finding: To measure the proportion of viable MLM in a suspension, inoculation of mice into the hind foot pad with serially diluted suspensions of the organisms was attempted, as has been done for M. leprae (25). The resulting multiplication of MLM after inoculation of small numbers of the organisms appeared to be much more rapid than the published estimates (24). Although Marchoux and Chorine had stated (9), as early as 1938, that as few as five organisms were sufficient to produce infection of the rat, no detailed study of the infection of mice produced by inoculation of small numbers of MLM into the foot pad has been reported.
Of considerable interest was the opportunity to examine an important aspect of the absence of an effective immune response to the organism, that is associated with MLM disease of mice of susceptible strains (3), particularly the question of whether the immune deficiency was a direct consequence of the infection, i.e., of multiplication of the organisms, or whether the large inocula usually employed in the course of experimental work with MLM were themselves immunosuppressive.
MATERIALS AND METHODS
Mice, 6-10 weeks of age, and of both sexes of two strains were employed: CBA/Ca, bred locally from a nucleus obtained approximately 3 years earlier from the National Institute for Medical Research (NIMR), London, U.K., and BALB/cABom, bred locally from a nucleus obtained at approximately the same time from Gl. Bomholtgard, Ry, Denmark. The mice were bred and housed under conventional but clean conditions. MLM Douglas strain were originally obtained from Dr. R. J. W. Rees, NIMR, and subsequently maintained in passage in CBA mice.
Inoculation of the mice and the subsequent harvest and enumeration of MLM were performed by methods described by Shepard for M. leprae (20,22). Inocula were prepared from the spleens and livers of mice that had been inoculated intraperitoneally 3 or 4 months earlier. MLM were enumerated in the resulting suspensions, diluted to a concentration of 5000 organisms per 30 µl, and a volume of the diluted suspension sufficient to distend fully the foot-pad tissues was inoculated, after which the injected volume was measured. At intervals after inoculation, harvests of MLM were performed from the inoculated foot pads, popliteal and inguinal lymph nodes, spleen, and liver. The tissues were minced with fine scissors and homogenized in a glass Ten Broeck tissue grinder (Bellco Glass Co., Vineland, New Jersey, U.S.A.), after which 10 µl of each suspension was transferred to the circles of a Reich counting slide (Bellco). Each aliquot was spread evenly with a platinum wire over the surface of a circle, after which the slides were air dried on a level board and fixed by exposure to Formalin fumes. The slides were stained by a room-temperature acid-fast stain. MLM were then enumerated by counting all of the organisms seen in 60 microscope fields at a magnification of ×1250, employing a research microscope equipped with a planachromat oil-immersion objective and Köhler illumination.
On the occasion of the harvests of MLM, the lymph nodes, spleen, and liver of the mice were excised and homogenized. Also, at intervals after inoculation the thickness of the inoculated feet was measured by means of a spring-loaded dial-gauge caliper (The Dyer Co., Inc., Lancaster, Pennsylvania, U.S.A.).
Because MLM can be cultivated only with difficulty in -free media (12,14), the proportion of viable organisms was measured by inoculation of mice. The method employed for determination of the proportion of viable organisms in each inoculum was based on the assumption that, given sufficient time, one viable organism will multiply to a number large enough (> 105) to be taken as evidence of multiplication, and therefore of viability (25). The MLM were diluted, and 5000, 500, 50 or 5 organisms were inoculated into the hind foot pads of naive mice. Four or more months after inoculation, MLM were harvested from individual foot pads, usually 10 foot pads per dilution, and the 50% infectious dose (ID50), here defined as the total number of organisms, both living and dead, required to infect half of the mice, was calculated by means of the Spearman-Kärber technique (5,21).
RESULTS
Multiplication of MLM. During the first 8 months after inoculation, CBA mice appeared grossly normal; thereafter, however, there was a noticeable decrease in weight, a few mice demonstrated paralysis of the hind legs, and some deaths were recorded. BALB/c mice demonstrated no obvious change of appearance during the entire year, with the exception of swelling of the inoculated hind feet. One year after inoculation, as shown in Table 1, the median value for the footthickness of infected BALB/c mice was 2.9 mm, a value significantly larger [p < 0.001, Mann-Whitney U-test (23)] than that, 1.8 mm, for the control mice. The 1.7 mm median thickness of the inoculated hind feet of the CBA mice did not differ significantly from that of the uninfected control mice.

Multiplication of the organisms in various organs during approximately 1 year following inoculation of 5000 MLM into both hind foot pads of CBA and BALB/c mice is shown in Tables 2 and 3. These experiments were not performed at the same time, nor with the same inoculum; in fact, the ID50 of the inoculum employed for the CBA mice was 3.16; whereas that for the inoculum employed for the BALB/c mice was 88.7. Thus, comparison of multiplication of the organisms in mice of the two strains at the early intervals is not justified. Nevertheless, both strains demonstrated a more rapid increase of bacterial numbers soon after inoculation, with a slowing of the rate of multiplication once the number of organisms per foot pad passed 3 x 107. The number of MLM per foot pad 1 year after inoculation was significantly [p < 0.05 (6)] larger in BALB/c than in CBA mice.
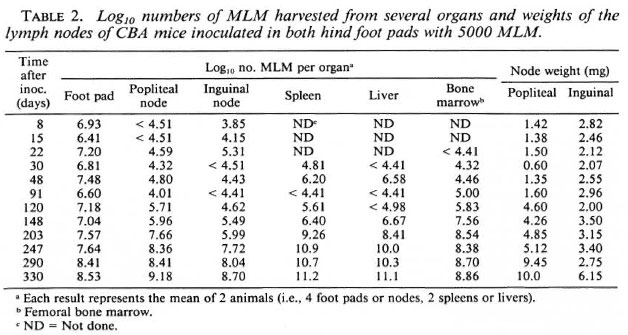
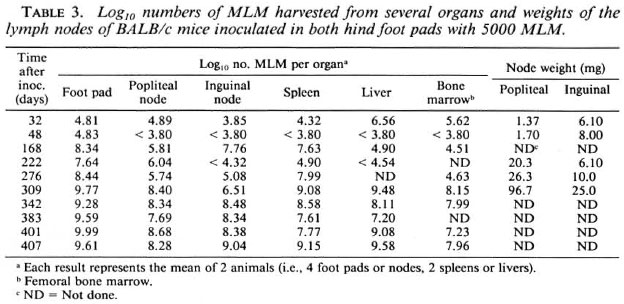
During the year following inoculation of CBA mice in the hind foot pad with 5000 MLM, the numbers of organisms (Table 2) were noted to reach levels > 1011 in the spleen and liver and >108 in the femoral bone marrow. As shown in Table 3, the numbers of MLM also reached high levels in the organs of BALB/c mice but not, at least in this experiment, the very large numbers found in CBA mice.
Differences between the CBA and BALB/c mice were also noted in terms of the popliteal and inguinal nodes, the lymph nodes primarily draining the foot pads (11). Among CBA mice (Table 2), beginning about 4 months after infection progressive increases of node weight and of the numbers of MLM recovered from the nodes are evident, despite considerable variation from animal to animal. Among BALB/c mice (Table 3), the numbers of MLM and the weights of the lymph nodes increased greatly over the course of the infection. Although the numbers of organisms harvested from the nodes were much the same in both strains of mice the increase of node-weight was much greater in the BALB/c strain than in the CBA mice.
In two additional experiments (Figs, 1a and 1b), CBA and BALB/c mice were inoculated into both hind foot pads with 5000 MLM per foot pad, the mice being inoculated with the same suspension, and harvests were performed at intervals of one to several days during the first 2 weeks after inoculation, and, in one of the experiments (Fig. 1b), at longer intervals thereafter. The growth curves suggest that, in fact, initial multiplication is similar in both strains of mice.
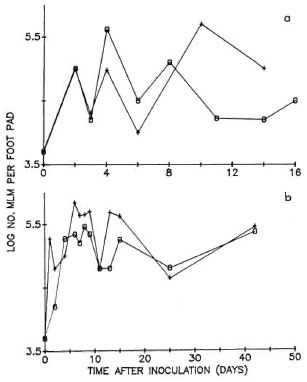
Fig. 1. Multiplication of MLM in hind foot padsof CBA (0) and BALB/c (+) mice after local inoculation of 5000 organisms. Every point represents the mean number of organisms harvested individually from four foot pads; a and b show the results of two experiments.
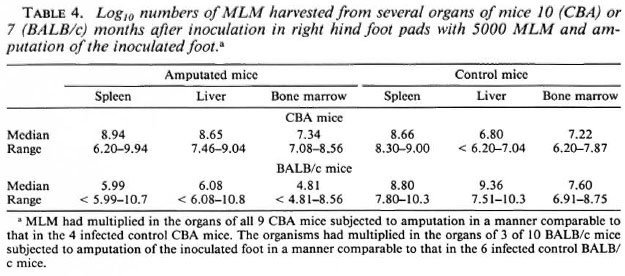
Dissemination versus multiplication in situ. The large numbers of MLM recovered from the spleens and livers of mice that had been inoculated months earlier with a small number of organisms in the hind foot pad raised the question of whether the large numbers represented dissemination from the site of inoculation or multiplication in situ. To answer this question, mice of both strains were inoculated in the right hind foot pad with 5000 MLM; 1 hr later, under pentobarbital anesthesia, the right foot was amputated at the ankle joint. Control mice were inoculated in the right hind foot pad but were not subjected to amputation. Seven months later for CBA mice, and 10 months after inoculation and amputation for BALB/c mice, MLM were harvested from the spleen, liver and femoral bone marrow of both amputated and control mice.
The results of these experiments (Table 4) indicate that in all of the 9 CBA mice and 3 of 10 BALB/c mice that had been inoculated in the right hind foot pad, and subjected to amputation of the right foot 1 hr later, MLM had multiplied in the spleen, liver and femoral bone marrow to large numbers, comparable to those encountered in the control animals.
Small inocula. The virulence of MLM for both BALB/c and CBA mice is demonstrated most clearly by the results of the experiments in which mice were inoculated in both hind foot pads with 5 or 50 organisms, after which the mice were observed for 1 year. By the end of the year, some deaths had occurred, and many of the surviving mice were sick. At this time, as shown in Table 5, harvests of MLM from the inoculated foot pads and spleens demonstrated that the organisms had multiplied to numbers > 108 in the foot pads, and to >; 109 in the spleens of most of the mice examined.
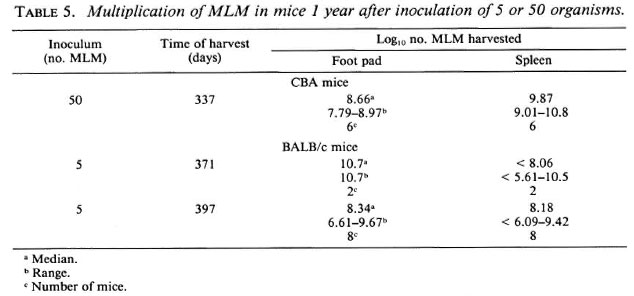
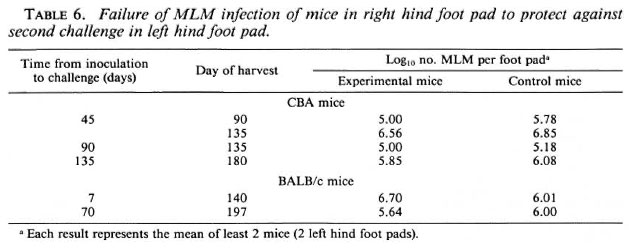
Response to a second challenge. To find an explanation for the virtually unlimited multiplication of the MLM after local inoculation of a small number of organisms, experiments were carried out to examine the response in both CBA and BALB/c mice to a second challenge with MLM. For this purpose, mice were inoculated in the right hind foot pad with 5000 MLM and were later inoculated, together with naive control mice, in the left hind foot pad with the same number of organisms. As shown in Table 6, no protection against the second challenge was demonstrated in CBA mice administered the second challenge 45, 90 or 135 days after the initial infection, nor were BALB/c mice protected against a second challenge administered 7 or 70 days after the initial infection.
Early killing. Because of the apparent decrease of the number of MLM per foot pad after initial multiplication in CBA mice (Fig. 1a), an additional experiment was performed, in the course of which the MLM harvested 21 days after inoculation were serially diluted and subinoculated into naive mice. The ID50 of the inoculum was determined to be 3.16; that of the organisms harvested 21 days later was 1253. Assuming that the ID50 during the initial phase of rapid multiplication was no smaller than that determined for the inoculum, the initial multiplication appears to have been followed immediately by death of at least 99% of the MLM.
To confirm early killing of the MLM, groups of CBA mice were inoculated into both hind foot pads with 5000 MLM per foot pad (ID50 3.16), and harvests were performed at close intervals thereafter. As shown in Figure 2, the number of MLM per foot pad increased rapidly during the first 24 hr. Subsequent harvests performed at intervals of 1 or 2 days demonstrated numbers of MLM ranging from 6300 to 6.3 × 105 per foot pad. Although the irregular multiplication depicted in this figure may simply represent animal-to-animal variation, the ID50 of the organisms harvested 5 days after inoculation was determined to be 125, indicating death of at least 97% of the organisms shortly after initial multiplication. Multiplication of MLM was observed to have resumed after day 16 but at a rate slower than that observed initially, reaching a value of 2.5 x 107 MLM per foot pad 90 days after inoculation.
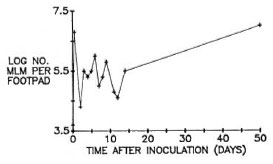
Fig. 2. Multiplication of MLM in hind foot pads of CBA mice after local inoculation of 5000 organisms. Early harvests were made at close intervals to demonstrate more clearly the decrease in number of organisms per foot pad soon after multiplication reached a maximum.
DISCUSSION
Subcutaneous infection of mice with MLM appears to be qualitatively different from subcutaneous infections of mice with other mycobacteria. Inoculation of mice in a hind foot pad with small numbers (e.g., 5000) of M. leprae or M. marinum results only in a self-limited process (13,20); whereas inoculation of larger numbers of organisms appears, particularly in the case of M. leprae, to provoke an immune response that prevents multiplication of the organisms (16). Virtually all of the published studies of subcutaneous MLM infection of mice have been carried out with inocula of 105 or more organisms, and the results of infection with 5000 MLM were not known. The possibility remained that mice, even those of susceptible strains, could limit multiplication of MLM if smaller numbers were inoculated.
The results of the present study demonstrate that multiplication of MLM to very large numbers occurs among susceptible mice that have been inoculated locally with small numbers of organisms. Moreover, despite the differences between mice of a more susceptible (BALB/c) and a less susceptible (CBA) strain (1), the organisms appear to have multiplied virtually without limit in the foot pads and lymph nodes of mice of both strains, and the infection underwent dissemination to approximately the same extent in mice of both strains. Thus, the progressive nature of MLM-infection of mice appears not to depend upon inoculum size, but, rather, to be a characteristic of the infection. This hypothesis finds confirmation in the multiplication of MLM to as many as 1010 per foot pad and spleen of mice that had been inoculated in the hind foot pad with as few as five organisms.
An interesting finding was the multiplication of MLM to very large numbers in the spleen, liver, and femoral bone marrow of mice that had been inoculated a number of months earlier with 5000 MLM in one hind foot pad, and had been subjected 1 hour later to amputation of the inoculated foot. Work in mice inoculated in a hind foot pad with 5000 M. leprae or M. marinum had demonstrated that 80% to 90% of the inoculum could not be recovered from the inoculated foot pad 1 hour after inoculation (8,10); only a small fraction of the missing organisms were located in the popliteal lymph nodes or spleen (10), suggesting that the organisms had entered the circulation and were widely distributed. Multiplication of MLM in the organs of the amputated mice indicates that MLM multiplied in situ. On the other hand, the more uniform multiplication in the organs of the control mice suggests that dissemination of MLM occurs as the organisms multiply in the inoculated hind foot pad and, perhaps also, in the draining lymph nodes.
Although we did not examine the immune response of the MLM-infected mice directly, one may infer from the very large numbers of MLM present in the foot pads and popliteal and inguinal lymph nodes 1 year after inoculation that the mice had been unable to limit multiplication of the MLM, even when only a small number of organisms had been inoculated subcutaneously. More direct evidence of immune unresponsiveness is the failure, demonstrated in this study, of an initial infection in one hind foot pad to protect mice against a second challenge in the contralateral foot pad. Mice infected in one hind foot pad with M. leprae or M. marinum evince solid resistance to a second challenge(13).
Although MLM-infected mice, even those infected locally with small numbers of the organisms, are unable to control the infection, probably because T-effector mechanisms are suppressed (2,15), the animals appear able to kill the organisms by other means that do not involve T lymphocytes. We showed earlier (17,18) that late during MLM-infection of mice, the majority of the MLM recovered from several organs of the mice were incapable of multiplying in mice and, therefore, were presumed to be dead. That this same phenomenon was encountered in nu/nu mice excluded T--mediated bactericidal mechanisms (17).
The mechanism by which large proportions of the organisms are killed early in the course of the process that follows local inoculation of small numbers of MLM is unclear. Rojas-Espinosa and his coworkers, who studied mice that had been inoculated with 108 MLM intraperitoneally, described (9) early, i.e., 45 days after inoculation, killing of MLM by macrophages, which they believed to have been activated by T-mediated mechanisms. We believe, however, that T--mediated activation of macrophages cannot account for early killing which, in the experiments described in this present report, appears to involve two phenomena occurring in mice within a few days after local inoculation of 5000 MLM: a) the decrease of the proportion of viable organisms demonstrated 21 days after inoculation in one experiment and 5 days after inoculation in another, and b) the abrupt decrease of the rate of multiplication that occurs once initial, rapid multiplication has carried the number of organisms to near 107 per foot pad. If these two phenomena are related, two arguments against T--mediated macrophage activation may be considered.
First, although we did not attempt to demonstrate early killing in nude mice, the growth curves of MLM in BALB/c nu/nu mice appeared virtually identical in form to those in BALB/c +/ + mice (F. Silbaq, et al., unpublished data). In particular, the organisms appeared to multiply rapidly to the level of 106 to 107 per foot pad within the first 1 or 2 days after inoculating the nude mice with 5000 MLM per foot pad, after which multiplication continued, but at a much slower rate as was shown to occur in immunologically competent CBA mice (24). If, in fact, early killing of MLM underlay the abrupt decrease of the rate at which MLM multiply in the foot pad, the bactericidal mechanism could not involve T lymphocytes.
The second argument against T--mediated activation of macrophages is that the abrupt decrease of the rate of multiplication occurs within 1 or 2 days following inoculation. This militates against a T--mediated mechanism; expansion of antigenspecific T- clones is not expected to occur so soon after first exposure to the antigen.
REFERENCES
1. BRETT , S. J. T-cell responsiveness in Mycobacterium lepraemurium infections in a "resistant" (CBA) and a "susceptible" (BALB/c) mouse strain. Cell. Immunol. 89(1984)132-143.
2. BROWN , I. N. Animal models and immune mech anisms in mycobacterial infection. In: Biology of the Mycobacteria, Vol. 2. Ratledge, C. and Stan ford, J., cds. New York: Academic Press, 1983, pp. 173-234.
3. BROWN, I. N., GLYNN, A. A. and PLANT, J. Inbred mouse strain resistance to Mycobacterium lepraemurium follows the Ity/Lsh pattern. Immunol. 47(1982)149-156.
4. CLOSS, O. Experimental murine leprosy; growth of Mycobacterium lepraemurium in C3H and C57BL mice after foot pad inoculation. Infect. Immun. 12(1975)480-489.
5. FINNEY, D. J. Statistical Method in Biological Assay. New York: Hafner Publishing Company, 1964, pp. 524-530.
6. KRUSHAT, W. M., SCHILLING, K. E., EDLAVITCH, S. A. and LEVY, L. Studies of the mouse foot-pad technique for cultivation of Mycobacterium leprae. 4. Statistical analysis of harvest data. Lepr. Rev. 47 275-286.
7. LEVY , L. Superinfection in mice previously infected with Mycobacterium leprae. Infect. Immun. 11(1975)1094-1099.
8. LEVY, L., M OON, N., M URRAY, L. P., O'NEILL, S. M., G USTAFSON , L. E. and E VANS, M. J. Studies of the mouse foot pad technic for cultivation of Mycobacterium leprae. 1. Fate of inoculated organisms. Int. J. Lepr. 42(1974)165-173.
9. MARCHOUX, E. and C HORINE, V. Cinq bacilles de Stefansky suffisent pour infecter le rat blanc. Ann. Inst. Pasteur 61(1938)291-299.
10. MOR, N. and LEVY, L. Importance of the footpad lesion in the mouse response to local inoculation of Mycobacterium marinum. Ann. Inst. Pasteur/ Microbiol. 136A(1985)191-201.
11. MOR, N., LUTSKY, I. and LEVY , L. Response in the distant lymph nodes of mice to infection in the hind foot pad with Mycobacterium marinum. Infect. Immun. 28(1980)225-229.
12. NAKAMURA, M. Quantitative multiplication of Mycobacterium lepraemurium in a -free liquid medium (NC-5). J. Gen. Microbiol. 82(1974)385-391.
13. NG, H., JACOBSEN, P. L. and LEVY , L. Analogy of Mycobacterium marinum disease to Mycobacterium leprae infection in footpads of mice. Infect. Immun. 8(1973)860-867.
14. PORTAELS, F. and P ATTYN , S. R. Parameters influencing the in vitro growth of Mycobacterium lepraemurium. Int. J. Lepr. 49(1981)194-197.
15. PTAK , W., G AUGAS , J. M., REES, R. J. W. and ALLISON, A. C. Immune responses in mice with murine leprosy. Clin. Exp. Immunol. 6(1970)117-124.
16. REES, R . J. W. Enhanced susceptibility of thymectomized and irradiated mice to infection with Mycobacterium leprae. Nature 211(1966)657-658.
17. RESNICK, M., BERCOVIER, H., AIZER, F., MOR , N. and LEVY, L. Death of Mycobacterium lepraemurium after multiplication in CBA, BALB/c and nude mice. Ann. Inst. Pasteur/Microbiol. 138(1987)15-19.
18. RESNICK, M., BERCOVIER, H., MOR , N. and LEVY, L. Unforeseen death of Mycobacterium lepraemurium in infected susceptible mice. J. Infect. Dis. 153(1986)368-370.
19. ROJAS-ESPINOSA, O., VEGA, R., OLTRA, A., ARCE, P. and NUNEZ, A. Transitory macrophage activation in the granulomatous lesions of Mycobacterium lapraemurium-induced lepromatoid leprosy in the mouse. Int. J. Lepr. 56(1988)428-436.
20. SHEPARD , C. C. The experimental disease that follows the injection of human leprosy bacilli into foot-pads of mice. J. Exp. Med. 112(1960)445-454.
21. SHEPARD , C. C. Statistical analysis of results obtained by two methods for testing drug activity against Mycobacterium leprae. Int. J. Lepr. 50(1982)96-101.
22. SHEPARD, C. C. and MCRAE, D. H. A method for counting acid-fast bacteria. Int. J. Lepr. 36(1968)78-82.
23. SIEGEL, S. Nonparametric Statistics for the Behavioral Sciences. New York: McGraw-Hill, 1956, pp. 116-127.
24. SILBAQ, F., LEVY, L. and BERCOVIER, H. Doubling time of Mycobacterium lepraemurium in the foot pads of mice. Res. Microbiol. 140(1989)43-50.
25. WELCH, T. M., GELBER, R. H., MURRAY, L. P., No, H., O'NEILL, S. M. and LEVY, L. Viability of Mycobacterium leprae after multiplication in mice. Infect. Immun. 30(1980)325-328.
1. M.Sc, Departments of Comparative Medicine and Clinical Microbiology, Hebrew University-Hadassah Medical School, P.O. Box 1172, 91010 Jerusalem, Israel.
2. Ph.D., Department of Comparative Medicine, Hebrew University-Hadassah Medical School, P.O. Box 1172, 91010 Jerusalem, Israel.
3. M.D., Ph.D., Department of Comparative Medicine, Hebrew University-Hadassah Medical School, P.O. Box 1172, 91010 Jerusalem, Israel.
4. D.V.M., Department of Clinical Microbiology, Hebrew University-Hadassah Medical School, P.O. Box 1172, 91010 Jerusalem, Israel.
Reprint requests to Prof. L. Levy, Department of Dermatology, Hadassah University Hospital, P.O. Box 12000, 91120 Jerusalem, Israel.
Submitted by F. Silbaq in partial fulfillment of the requirements for the M.Sc. degree.
Received for publication on 4 December 1989.
Accepted for publication in revised form on 5 June 1990.