- Volume 57 , Number 4
- Page: 874–9
Cultivation of Mycobacterium leprae: a new approach
To the Editor:
We wish to report our recent findings on our Mycobacterium leprae culture isolates-ICRC strains which may throw new light on the problem of cultivation of M. leprae in vitro and on the origin of leprosy-derived mycobacteria (LDM). In the course of our investigation on antigenic differences among M. leprae culture isolates and on the development of an improved vaccine, we discovered that the ICRC bacilli are not pure cultures, but consist of two mycobacteria with distinctive properties.
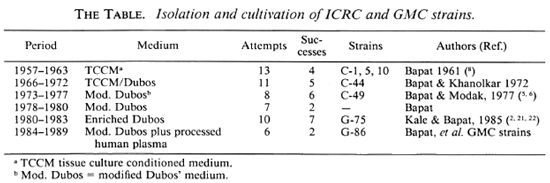
Since 1958, in our attempts to grow M. leprae in vitro, we have isolated several strains of mycobacteria designated as ICRC bacilli (7,8). These cultures have been grown exclusively on liquid media, e.g., tissue culture conditioned medium and enriched Duces' medium (8,21) (The Table). On the other hand, the majority of LDM are grown on solid media, and are found to belong to the M. avium-intracellulare-scrofulaceum (MAIS) complex. Since some of the ICRC strains isolated during 1958-1961 had shown lepromin-like reactivity, the only marker of M. leprae identification available at the time, we continued the work on new isolates with detailed studies on some of the strains (The Table). They were found to share some M. leprae-specific characteristics, e.g., lepromin-like activity (6), growth in the mouse foot pad (5), DOPA-oxidase activity (22), and antigenic behavior like that of M. leprae in mice (1,20). However, they express a biochemical profile like that of M. avium-intracellulare (8,21 and Kato, L., personal communication, 1979). A candidate antileprosy vaccine prepared from one of the strains was found to induce lepromin conversion and reversal reactions in LL patients, and has the efficacy and potency equivalent to that of BCG plus M. leprae vaccine (10,12,13 and Bapat, et al, personal communication, 1980). Experimentally, the irradiated ICRC bacilli were found to induce strain-specific sensitization in mice (20). A similar pattern of sensitization, i.e., differential sensitization to different ICRC strains, was observed in both ICRC-vacci-nated LL patients and unvaccinated lepro-matous patients (4). The observations summarized above strongly indicated the apparent presence of "M. leprae" activity in the ICRC cultures.
The nature of M. leprae culture isolates-ICRC bacilli has remained enigmatic for a long time, particularly because of the expression of dual characteristics of both M. leprae and M. avium-intracellulare. A possibility of "recombination" or "fusion" of two mycobacteria as a mechanism for the phenomenon has been proposed (2). However, the fact that the two sets of enzyme characteristics are mutually exclusive indicates that the mechanism may be remote. An alternate hypothesis of the existence of two mycobacteria in the ICRC culture was presented in 1986 (3). This concept of two populations appeared to be a better and more practical approach to explore the issue.
We already know that an inoculum of ICRC bacilli on Löwenstein-Jensen (LJ) medium yields M. avium-intracellulare (8 and Kato, L., personal communication, 1979). On this background, we preferred and adopted a physical method-density gradient centrifugation (19) -to search for the other component, if any, in the ICRC culture harvest. The results of the experiments revealed that the long-term M. leprae culture isolates-ICRC bacilli did, indeed, contain just two distinct organisms. This discovery, that the ICRC culture is not a pure strain but consists of two mycobacteria, thus supports the hypothesis (3). By connotation, the discovery has considerable significance and importance for the cultivation problem and the vaccine design. Therefore, we examined the two fractions for M. leprae-specific markers. The data collected are presented in this communication, mentioning a few immediate implications.
MATERIALS AND METHODS
Cultures. The ICRC strains (recalled from Dr. L. Kato, Catherine Booth Hospital Centre, Quebec, Canada) and the Grant Medical College (GMC) strains-new isolates-were grown in modified Dubos' medium with human serum (21). A type-strain M. avium was grown in Sauton's medium. A few samples of fresh M. leprae were prepared from human LL skin biopsies collected aseptically.
Gradient system. A discontinuous sucrose density gradient (5%-30%) was made and approximately 1010 organisms were loaded immediately on it. The tubes were ccntrifuged at 1100 x g x 20 min. The results showed a clear-cut dissociation of the original suspension into two components, the "light" band of 1.07 g/ml density and the pellet of "heavy" organisms of > 1.12 g/ml density. Further enrichment was done routinely. Under similar conditions, on a discontinuous Percoll (19,27) gradient, the "light" population separated at the density of 1.06 g/ml; the "heavy" population formed a sharp band at the density of 1.09 g/ml. The separation was slow (1 hr 20 min), but the bands were very sharp and stable (The Figure). The results of separation were reproducible with all the strains currently maintained as well as with stored samples.
The Figure. Separation of "light" and "heavy" populations on density gradients at 1100 x g. A = Sucrose, 20 min; B = Percoll, 80 min.
Identification tests. The primary aim was to establish that they were two different mycobacteria. The studies were conducted on two strains, grown in large quantity, and the fractions were subjected to the tests adopted for M. leprae-specific markers.
OBSERVATIONS
Properties of "light" and "heavy" organisms
The yield. The proportionate yield of the "light" and the "heavy" organisms was 30:70 on a wet weight basis of the total of 1 g/1 at 3 weeks.
Morphology and staining characteristics. Both of the fractions contained acid-fast rods, the majority of which were solidly stained. The "light" acid-fast bacilli (AFB) were slender with uniform spreading; the "heavy" AFB were thicker with few clumps.
Pyridine extraction (11) and Ziehl-Neel-sen staining. After a 2-hr extraction with fresh pyridine, a slide preparation of the "light" and the "heavy" organisms, along with a fresh M. leprae smear as the control, indicated that the "light" AFB lost acid-fastness (also M. leprae); the "heavy" AFB did not do so and showed pink rods.
D-DOPA oxidase activity (25). The results were reproducible in all of the tests with standardized suspensions, showing that the "light" organisms readily oxidized D-DOPA. They even showed pigment formation on the sediment. The experiments showed ΔOD (480 nm) = 0.16-0.2; the activity of the "heavy" AFB was ΔOD 0.03-0.06 as that of the heat-killed bacilli.
Catalase activity. Catalase activity was negative for the "light" AFB and positive for the "heavy" AFB.
Aryl-sulfatase. The results were positive for the "light" AFB and negative for the "heavy" AFB.
Tellurite reduction. Tellurite reduction was negative for the "light" organisms; positive for the "heavy" bacilli.
Detection of PGL-I antigen (16). An ELISA with polyclonal rabbit antibody to phenolic glycolipid-I (PGL-I) showed a higher concentration of PGL-I on "light" AFB than on the "heavy" AFB.
Growth on LJ medium. The inoculum of "light" organisms did not show growth at 4 weeks; in the same period, the "heavy" AFB produced small colonics of AFB.
Further studies on purified "light" and "heavy" AFB for their lepromin activity, extraction of mycoside, genome size, G + C content, DNA homology, protein profiles of sonicates by SDS-PAGE and by the Amibs protein method are in progress.
Type strain M. avium
M. avium bacilli grown in Sauton's medium and placed on the sucrose gradient were found to contain only the "heavy" organisms (D = 1.12 g/ml) and no diffuse distribution of any lighter organisms.
Study of fresh M. leprae suspension from human skin biopsy. The reproducibility of fractionation with small samples of M. leprae culture isolates prompted us to examine the fresh, human M. leprae suspension. A 3-ml suspension of minced tissue was filtered through thin cotton mesh, and the filtrate was loaded on the gradient. The organisms separated as "light" AFB (D = 1.07) and "heavy" AFB. The "light" AFB in the smear contained many AFB. the majority were beaded and a few solidly stained (original bacterial index 5 +), and showed total loss of acid-fastness on the pyridine extraction (PE) test. The "heavy" AFB were few in number and were found to be PE resistant. The paucity of the material and scarcity of the LL biopsies prevented any other tests, except for the inoculation onto modified Dubos' medium.
COMMENTS
The above data show that M. leprae iso-lates-ICRC strains contain two distinct mycobacteria, and this appears to be the characteristic of all (ICRC and GMC) strains grown in liquid media. The evidence supports the concept of two populations as presented in 1986 (3). The properties of the "light" and the "heavy" AFB examined so far establish that they are two distinct mycobacteria, both physically and bacteriologically. They grow as an established co-culture system, and the dissociation of the two mycobacteria has been demonstrated for the first time.
The "heavy" AFB, the major component from available data, correspond to At. avium-intracellulare. The "light" AFB sharing M. leprae characteristics, particularly the D-DOPA-oxidase activity at the culture level, may correspond to the "M. leprae activity" shown by the ICRC bacilli (22). The final identity status of the "light" AFB will be established when DNA homology data are available.
We believe that detection of a small number of PE-resistant "heavy" organisms in a fresh M. leprae suspension may be the key to the question of the origin of LDM. In almost all of the attempts to grow M. leprae in culture, when growth is seen it is invariably of mycobacteria belonging to the MAIS complex, irrespective of the medium, the source of M. leprae, or the heavy size of inoculum (15,26). Their presence in the original inocula has been generally accepted (15,26). From our observations, it is not difficult to imagine a few "trapped" "heavy" organisms in purified M. leprae preparations from armadillo-liver tissue which may be the seed of LDM. The AFB in the sediment obtained during M. leprae (A) purification appears to have been ignored. The survival and limited growth of M. leprae in a variety of special media has been reported with the evidence of growth in the mouse foot pad, but eventually they seem to be swamped by MAIS, and/or eliminated through subcultures (14,24,26).
Kato (23) had proposed that "concomitant mycobacteria play a role of an etiological co-factor by providing mycobactin to promote the growth of M. leprae in vivo (17,23).Based on this principle, he has placed M. leprae in the category of a microbe-dependent microorganism of the M. paratuber-culosis type. We seem to have empirically developed a similar system with the ICRC culture, unknowingly using the same concept. This constant yield of "light" bacilli with a growth limit at 3 weeks suggests a mutually controlled growth of the two mycobacteria (8,15).
The "light" AFB obtained from M. leprae culture isolates of the ICRC type, in all probability, could be very close to M. leprae. This could have been the component capable of growth in the mouse foot pad (5); the source of protein antigens crossreacting with M. leprae-derived proteins (9); and the organisms with epitopes reacting with human T-cells derived from TT patients (18). The responses induced by the ICRC vaccine in LL patients, particularly the reversal reaction mentioned earlier, indicate that the "light" AFB component may be antigeni-cally closer to M. leprae than any other my-cobacterium. This revelation, that the ICRC bacilli contain an AFB component sharing M. leprae antigens mixed with M. avium-intracellulare, explains why a single injection of the ICRC vaccine is as effective as Convit's M. leprae plus BCG vaccine. The ICRC vaccine preparation inherently embodies exactly the same rationale on which Convit designed his vaccine.
The "light" AFB component appears unable to grow as a single entity. However, the co-culture method and subsequent separation offers a rapid quantitative assay to measure the growth and to evolve a better medium for higher yields of the "light" AFB with other mycobacteria or growth factor such as mycobactin. The practical applications of the system are many: a) a quality control standard to monitor the "light" AFB content of the vaccine batch and to increase potency; b) isolation of pure organisms from a single biopsy source against pooled bacilli leading to the study of strain variations with reference to antigenic epitopes; c) design of a polyvalent vaccine by combination of the "light" AFB from the different strains, e.g., derived from drug-resistant M. leprae; and d) to answer many unsolved questions.
In brief, we believe that through the co-culture system yielding ICRC/GMC strains from M. leprae isolates a new avenue is opened up to have a fresh look at the cultivation problem. The culture method has been reproducible for many years. It is expected that this will pave the way to solve the most formidable, century-old puzzle in microbiology, namely, the cultivation of M. leprae in vitro.
- Dr. C. V. Bapat
Emeritus Medical Scientist (ICMR)
% Neuropathology Unit
Grant Medical College
Bombay 400 008, India
Acknowledgments. The project on leprosy has been sponsored by the Maharashtra Council for Science and Technology, Government of Maharashtra; Birla Smar-ak Kosh (BSK), Bombay Hospital: and the Indian Council of Medical Research (ICMR), New Delhi, India. The laboratory facilities were provided by Grant Medical College (GMC), Bombay. The experiments were conducted by Dr.(Mrs.) Vaijayanti P. Kale and Miss Aarti Trivedi supported by BSK, and by Miss Sheela Nair. Grateful thanks are due to Dr. K. B. Sainis, BARC; Dr. P. R. Mahadevan, Foundation for Medical Research, and Dr. W. S. Bhatki and Mr. R. G. Chu-lawala of Acworth Leprosy Hospital; Dr. Padhi, T.I.F.R., and Dr. Damayanti Shah, R.M.C. M. leprae (A), lepromin, and soluble M. leprae sonicate were kindly supplied by Dr. R. J. W. Rees, Medical Research Council, London.
REFERENCES
1. Amminikutty, J., Bapat, C. V. and Deo, M. G. Adoptive transfer of tolerance induced by ICRC bacilli against Mycobacterium leprae in mice. Int. J. Lepr. 54(1986)437-445.
2. Bapat, C. V. Immunological properties of M. leprae culture isolates ICRC bacilli: hypothesis on relationship between M. leprae and ML-culture isolates. Acta Leprol. (Geneve) 2(1984)175-194.
3. Bapat, C. V. The ICRC-culture strains derived from M. leprae: their significance to cultivation of M. leprae and leprosy spectrum. In: Proceedings of the Indo-U.K. Symposium on Leprosy, Agra, April 7-10, 19S6. Agra: The Coronation Press, 1987, pp. 250-253.
4. Bapat, C. V. and Kale, V. P. Differential sensitivity to ICRC strains by LMI tests in vaccinated and unvaccinated leprosy patients and in mice; its significance to antileprosy vaccine. Abstract P-32. 4th International Colloquium on Mycobacteria- Structure and Function. Pasteur Institute, Paris, September 1988.
5. Bapat, C. V. and Modak, M. S. Growth of ICRC bacilli in foot-pad of mice. Lepr. India 50(1978)144-155.
6. Bapat, C. V., Modak, M. S., D'Souza, N. G. A. and Chulawala, R. G. Immunological identification of ICRC-bacilli cultivated "in vitro" from M. leprae isolates by lepromin reactivity in leprosy patients. Lepr. India 49(1977)472-484.
7. Bapat, C. V., Randive, K. J. and Khanolkar, V. R. "In vitro" cultivation of an acid-fast mycobacterium from human lepromatous leprosy. Indian J. Pathol. Bacteriol. 1(1958)156-159.
8. Bapat, C. V., Ranadive. K. J. and Khanolkar, V. R. Growth characteristic studies on acid-fast mycobactcrium isolated from human lepromatous leprosy. Int. J. Lepr. 29(1961)329-342.
9. Chirmule, N. B., Mulherkar, R. and Deo, M. G. Antigenic profile of ICRC bacilli with special reference to isolation of immunogenic subunit. Int. Arch. Allergy Appl. Immunol. 86(1988)19-27.
10. Convit, j., Aranzazu, N., Ulrich, M., Zuniga, M., df. Aragon, M. E., Alvarado, J. and Reyes, O. Investigations related to the development of a leprosy vaccine. Int. J. Lepr. 51(1983)531-539.
11. Convit, J. and Pinardi, M. E. A simple method for differentiation of Mycobacterium leprae from other mycobacteria through routine staining techniques. Int. J. Lepr. 40(1972)130-132.
12. Deo, M. G., Bapat, C. V., Bhalerao, V., Chaturvedi, R. M., Bhatki, W. S. and Chula-wala, R. G. Antileprosy potential of ICRC vaccine; a study in patients and healthy volunteers. Int. J. Lepr. 51(1983)540-549.
13. Deo. M. G., Bapat, C. V., Chulawala, R. G. and Bhatki, W. S. Potential antilcprosy vaccine from killed ICRC bacilli: a clinico-pathological study. Indian J. Med. Res. 74(1981)164-177.
14. Dhople, A. M., Green, K. J. and Osborne, L. J. Limited "in vitro" multiplication of Mycobacterium leprae. Int. J. Lepr. 57Suppl.(1989)320-321.
15. Dhople, A. M. and Osborne, L. J. Presence of mycobactin-like substance in Mycobacterium leprae. Indian J. Lepr. 60(1988)348-359.
16. Douglas, J. T., Naka, S. O. and Lee, S. W. Development of an ELISA for detection of antibody in leprosy. Int. J. Lepr. 52(1984)19-25.
17. Draper, P. and Wheeler, P. R. Workshop on microbiology, XIII International Leprosy Congress. Int. J. Lepr. 57Suppl.(1989)301-302.
18. Emmrich, F. and Kaufmann, S. H. E. Human T-cell clones with reactivity to Mycobacterium leprae as tools for the charcterization of potential vaccines against leprosy. Infect. Immun. 51(1986)879-883.
19. Isolation of bacteria from yoghurt on gradients of Percoll® generated in situ. Standard Course Experiment. Uppsala: Pharmacia, 1982, p. 44.
20. Kale, V. P. and Bapat, C. V. Antigenic cross-reactivity between ICRC bacilli and M. leprae: an "in vitro" evaluation. Indian J. Lepr. 56(1984)219-231.
21. Kale, V. P., Bhat, A. V. and Bapat, C. V. Comparison of biochemical characters of ICRC bacilli with M. leprae: effect of substrate alteration in the medium. Indian J. Lepr. 56(1984)212-218.
22. Kale, V. P., Bhat, A. V. and Bapat, C. V. DOPA-Oxidase activity of ICRC bacilli. Indian J. Lepr. 56(1984)58-62.
23. Kato, L. Absence of mycobactin in Mycobacterium leprae: probably microbe dependent microorganism: implications. Indian J. Lepr. 57(1985)58-70.
24. Khera. V., Sawant, S. and Mahadevan, P. R. "In vitro" grown M. leprae-cultivablebacteria with conditioned phenotypic expression. Indian J. Lepr. 56 Special Issue (1984) abstract no. 223.
25. Prabhakaran, K., Harris, E. B. and Kirchhei-mer, W. F. O-Diphenoloxidase of Mycobacterium leprae separated from infected armadillo tissues. Infect. Immun. 12(1975)267-269.
26. Wheeler, P. R. Metabolism in Mycobacterium leprae: its relation to other research on M. leprae and to aspects of metabolism in other mycobacteria and intracellular parasites. (Editorial) Int. J. Lepr. 51(1984)208-230.
27. World Health Organization. Purification of M. leprae. WHO/TDR/IMMLEP/SWG(5)/80.3 (1980) p. 26.