- Volume 58 , Number 1
- Page: 44–9
Route-related variation in immunogenicity of mycobacteria
ABSTRACT
The route of immunization was observed to play a significant role in deciding the T-cell response to immunization with killed mycobacterial vaccines. Slow-growing mycobacteria were found to be immunogenic by both the intraperitoneal (i.p.) and intradermal (i.d.) routes; rapid-growing mycobacteria were immunogenic by the i.d. route only.
The nonresponder state following i.p. immunization with Mycobacterium vaccae could be corrected by treatment of the mice with poly 1: poly C or indomethacin prior to immunization. Both poly I: poly C, an interferon inducer, and indomethacin, a prostaglandin inhibitor, arc known to enhance the expression of major histocompatibility complex glycoproteins. Since they are so important in antigen preparation, it was concluded that the inability of mice to respond to M. vaccae by the i.p. route is likely due to defective presentation of the bacterial antigens by the antigen-presenting cells at the site, namely, the peritoneal macrophages. These findings are significant because M. leprae has been reported to be antigenically similar to M. vaccae, and the response of mice to i.p. immunization with both of these mycobacteria is very similar.
RÉSUMÉ
On a observé que la voie choisie pour l'immunisation joue un role important pour decider de la réponse des cellules-T à cette immunisation par des vaccins bactériens tués. Les mycobactéries à croissance lente sont immunogènes tant par la voie intrapéritéonéale (i.p.) que par la voie intradermique (i.d.). Les mycobactéries à croissance rapide ne sont immunogènes que par la voie i.d.
L'état de non réponse constaté à la suite de l'immunisation i.p. par Mycobacterium vaccae peut être corrigé par le traitement avant immunisation des souris par le poly I : poly C ou par l'indomédacine. On sait que le poly I : poly C, qui est un inducteur de l'interferon, de même que i'indomethacine, qui est un inhibiteur des prostaglandines. amplifient l'expression des glycoprotéines du complexe majeur d'histocompatibilité. Vu que les cellules qui présentent l'antigène, à savoir les macrophages péritéonéaux, jouent un rôle tellement important pour la présentation de l'antigène, on en a conclu que l'incapacité des souris à répondre à M. vaccae lors d'une immunisation par voie i.p., est vraisemblablement due à une présentation defective des antigènes bactériens par les macrophages. Ces observations sont importantes, car on a signalé que M. leprae était antigéniquement similaire à M. vaccae, et par ailleurs la réponse des souris à l'immunisation i.p. par l'un et l'autre de ces types de mycobactéries est très semblable.
RESUMEN
Se observó que la ruta de inmunización juega un papel importante en el resultado de la inmunización con vacunas micobacterianas muertas. Ya antes reportamos que las micobacterias de crecimiento lento fueron inmunogénicas tanto por la via intraperitoneal (i.p.) como por la vía intradérmica (i.d.). En contraste, las micobacterias de crecimiento rápido solo fueron inmunogénicas por la vía intradérmica. Ambos tipos de micobacterias generaron la clásica supresión mediada por células T Lyt-2 positivas después de la inmunización i.p. pero no después de la inmunización i.d. En el caso de micobacterias de crecimiento lento, la supresión mediada por LcT fue solo un componente de la respuesta generada después de la inmunización i.p. En contraste, en el caso de Mycobacterium vaccae y las otras de crecimiento rápido, la supresión mediada por células T fue la respuesta predominante a la inmunización i.p. La supresión mediada por células T generada por inmunización i.p. exhibió rectivicad cruzada, el espectro de la cual fue dependiente de la dosis de inmunización.
Leprosy and tuberculosis are characterized by phases of illness associated with immunological nonrespondcr states (5, 7). The pathogenesis of the phenomenon is not clear, but some have suggested a genetic defect in antigen presentation linked to major histocompatibility complex (MHC) encoded DR antigens (14, 15) From data obtained by the immunizing of mice with Mycobacterium leprae by different routes, others (8) have suggested that the pathogenesis of the nonrespondcr state could be related to the route of infection.
Using cultivable mycobacteria, we have studied the role of the routes of immunization on their immunogenicity. Also, in a separate publication (9) we have shown that the observed route-related variation in immunogenicity is due to variation in the presentation efficiency of antigen-presenting cells (APCs) encountering the antigen.
MATERIALS AND METHODS
M. tuberculosis H37Rv (NCTC 7416), M. kansasii (photochromogen, NCTC 10268), M. avium (nonchromogen, NCTC 8559), and M. phlei (rapid grower, NCTC 8151) were purchased from the National Collection of Type Cultures, London. M. vaccae (rapid grower) and M. scrofulaceum (scotochromogen) were kindly supplied by Dr. J. L. Stanford of Middlesex Hospital, London.
The mice used, the preparation of the sonic extracts, the immunizations, testing for delayed-type hypersensitivity (DTH), and the lymphoproliferation test (LT) were the same as described in the accompanying paper (10).
An IFN assay was carried out on the sera of mice treated with poly I: poly C by employing the technique described by Merigan (6). The results are expressed as International units--IU/ml - employing commercial gamma-IFN (Sigma, -I, 6507) as a standard.
RESULTS
Immunogenicity of mycobacteria by i.p. and i.d. routes of immunization. Swiss white mice were immunized with vaccines of different killed mycobacteria, in doses varying from 105 to 109 bacilli per mouse, suspended in 0.5 ml phosphate buffered saline (PBS) for intraperitoneal (i.p.) immunization and 0.05 ml PBS for intradermal (i.d.) immunization. The DTH response against the sonicate of the immunizing mycobacterium was assayed by the mouse foot-pad swelling technique (Table 1).
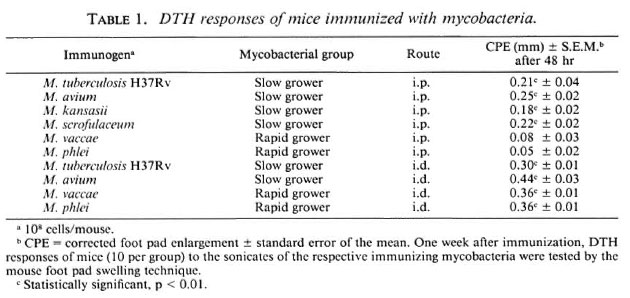
Slow-growing mycobacteria, namely, M. tuberculosis H37Rv, M. avium, M. kansasii, and M. scrofulaceum, generated a good DTH response on immunization by both i.p. and i.d. routes. The minimum immunogenic dose per mouse was 106 bacilli by both routes. The rapid growers, however, showed a totally different picture. M. vaccae and M. phlei failed to generate a DTH response on i.p. immunization, even at a dose of 109 bacilli per mouse. On the other hand, both of these organisms were immunogenic by the i.d. route, and the minimum immunogenic dose again was 106 bacilli per mouse.
The immunogenicity of M. tuberculosis, M. avium, and M. vaccaeby the i.d. and i.p. routes of immunization was also tested in the inbred BALB/c mice, and the results were identical. It was concluded that slow growers were immunogenic by both i.p. and i.d. routes; the rapid growers were immunogenic by the i.d. route only.
Spectrum of DTH crossreactivity. Swiss white mice were immunized with killed M. tuberculosis H37Rv (i.p. route) and M. vaccae (i.d. route) as before, and the DTH responses to the sonicates of several mycobacteria were tested (Table 2). In each mouse, one foot pad received the sonicate ofthe immunizing mycobacterium and the other foot pad received the sonicate of the mycobacterium for the testing ofcrossreactivity. Crossreactivity was observed at all immunogenic doses, and a similar crossreactivity spectrum was observed with other slow and rapid growers used as immunogens.
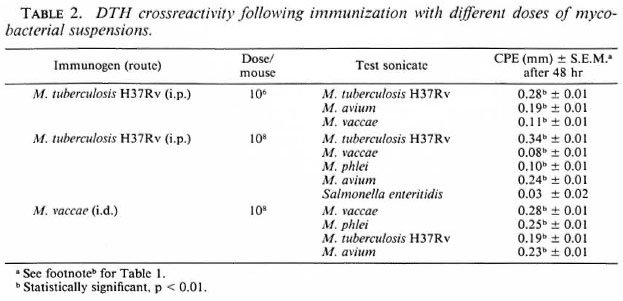
Effect of IFN inducer and indomethacin. Swiss white mice were treate travenous (i.v.) injection of 100 μg of poly I: poly C (an IFN inducer) 3 hr before receiving an i.p. immunization of 108 M. vaccae. One week later the DTH response to its sonicate was tested. A separate batch of 10 mice, similarly treated with poly I: poly C, were bled 3 hr after treatment, and the scrum IFN levels were assayed. The dose and time schedules for the poly I: poly C and indomethacin treatment were decided as per published reports(2,11). The mice showed a mean IFN level of 906 ± 371 IU/ml. It was observed that poly I: poly C-treated mice mounted a definite DTH response after i.p. immunization with M. vaccae; whereas the nontreated mice did not (Table 3).

The effect of indomethacin, a prostaglandin inhibitor, on i.p. immunization with M. vaccae was also investigated. Swiss white mice were given six i.p. injections of indomethacin (50 μ/mouse) at 12hr intervals. Twelve hours after the last dose, they were immunized with 108M. vaccae by the i.p. route and their DTH responses were tested. The indomethacintreated mice showed a DTH response; the nontreated micedid not. Theeffectsofthe poly I: poly C and indomethacin treatments on i.p. immunization with the other rapid growers were also tested and found to be identical.
In vitro lymphoproliferation test (LT) on spleen cells. Mice were immunized with 108 M. tuberculosis H37Rv or M. vaccae by the i.p. and i.d. routes. Their spleen cells were harvested and a LT was performed with 10 μg ofthe sonicate of the respective organism. As can be seen in The Figure, the spleen cells of those mice primed with M. tuberculosis H37Rv (by either the i.p. or i.d. route) gave a good in vitro proliferative response, not only to the sonicate of the immunizing organism but also to the sonicate of M. vaccae, which was shown earlier to crossreact in a DTH response. As expected, the proliferative response to the crossreacting M. vaccae sonicate was nowhere comparable to the proliferative response to the sonicate of the immunizing organism.
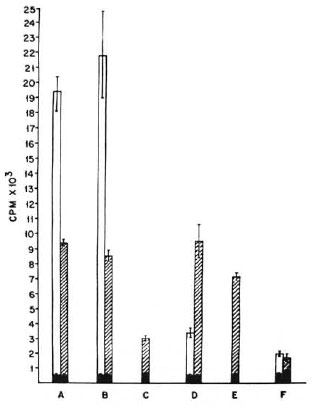
The Figure. Lymphoproliferative responses of spleen cells of BALB/c mice primed: A = i.p. with M.tuberculosis H37Rv; B = id. with M. tuberculosis H37Rv; C = i.p. with M. vaccae; D = i.d. with M.vaccae; E = i.p. with M. vaccae after 2 doses of poly I: poly C 24 hr apart; F = normal nonimmune BALB/c mice. CPM = mean count per min of 5 culture wells ± standard error of the mean (as bars).  = no antigen;
= no antigen;  = M. tuberculosis H37Rv sonicate;
= M. tuberculosis H37Rv sonicate;  = M. vaccae sonicate (10 µg/culture).
= M. vaccae sonicate (10 µg/culture).
The proliferative responses of the spleen cells of mice primed with M. vaccae (by the i.p. and i.d. routes) showed distinct differences. The spleen cells of i.d.primed mice showed a good proliferative response to M. vaccae and the crossreacting M. tuberculosis sonicate. In contrast, the proliferative responses of the i.p.primed spleen cells against both the immunizing M. vaccae and the crossreacting M. tuberculosis sonicates was very poor.
Thus, the lymphoproliferative responses of the i.p.and i.d.primed spleen cells showed close agreement with the in vivo DTH responses ofthe mice to the two immunogens given by the two routes.
DISCUSSION
This investigation was undertaken to evaluate the role of the route of immunization in mycobacterial infections. The ability ofdifferent killed mycobacterial vaccines to generate Tcell responses on immunization by the i.p. and i.d. routes was investigated in both outbred Swiss white and inbred BALB/c mice. For the study of in vitro antigenspecific lymphoproliferation, it was essential to pool the cells of several mice, which could only be undertaken in the inbred strains. Hence, both inbred and outbred mouse strains were employed.
Employing the in vivo DTH response and the in vitro lymphoproliferative response, it was shown that M. tuberculosis H37Rv and the other slow growers were immunogenic by both i.p. and i.d. routes ofimmunization. In contrast, M. vaccae and the other rapid growers were found to be immunogenic by the i.d. route only. Further, the nonresponder state of mice to the i.p. immunization with M. vaccae could be corrected by treatment of the mice with poly I: poly C, and IFN inducer (2), or indomethacin, a prostaglandin inhibitor, given before immunization.
One of the major factors influencing the outcome of immunization is the antigenpresenting cell (APC). Theoretically, the observed differences in the immunogenicity of M. tuberculosis H37Rv and M. vaccae when given by the i.p. route and the differences in the immunogenicity of M. vaccae when given by the i.p. and i.d. routes could be entirely accounted for by the differences in the efficiency of presentation of the two antigens by the APCs encountering the antigens, namely, Langerhans' cells and peritoneal macrophages (MΦ) (3, 4). Distinct differences in the density of major histocompatibility complex (MHC) class II or Ia glycoproteins that are so important in antigen presentation have been reported in these two types of APCs, being especially low in the latter (1, 3, 4). Langerhans' cells, being rich in Ia antigens, are generally expected to present antigens efficiently. On the other hand, with APCs poor in Ia density, such as the peritoneal MΦ, only those antigens that interact effectively with Ia glycoproteins would be expected to be presented with requisite efficiency for the generation of a Tcell response. Hence, the observed difference in the immunogenicity of M. tuberculosis and M. vaccae, when given by the i.p. route, could be an expression of poor presentation of M. vaccae antigen by the peritoneal cells. This is further supported by the fact that the nonresponder state to i.p. immunization by M. vaccae could be corrected by prior treatment with the prostaglandin inhibitor indomethacin and the IFN inducer poly I: poly C. Both of these agents are known to enhance Ia expression (11, 13). Thus, the data indirectly assign an important role to antigen presentation, the APCs, and their Ia composition in the pathogenesis of nonresponder states in mycobacterial infections. It may be argued that the in vivo effect of poly I: poly C and indomethacin could be quite complex and not restricted to peritoneal MΦ alone. In a separate publication (9), we present conclusive evidence to show that peritoneal MΦ are poor in presentation of M. vaccae antigen, which can be improved by treatment with poly I: poly C.
The data pertaining to the immunogenicity of M. vaccae are ofparticular significance because ofthe reported antigenic similarity of the organism to M. leprae (12,16). It may be noted that the immunogenicity of M. leprae to mice is very similar, i.e., it is immunogenic by the i.d. route only (8).
Acknowledgment. K. E. Shroff thanks the University Grants Commission, Government of India, for a fellowship grant to undertake this study.
REFERENCES
1. BELLER, D. I.. KIELY, J. M. and UNANUE, E. R. Regulation of macrophage population. I. Preferential induction of Ia rich peritoneal exudate by immunological stimuli. J. Immunol. 124(1980)1426-1432.
2. BRENAN. M. and ZINKERNAGEL, R. M. Influence of one virus infection on a second concurrent primary in vivo antiviral cytotoxic T-cell response. Infect. Immun. 41(1983)470-475.
3. ERB, P., RAMILA. G.. STUDER, S., LOEFFLER. H., CECKA, J. M., CONSCIENCE, J. F. and FELDMAN, M. Heterogeneity of accessory cells. Immunobiology 168(1984)141-153.
4. KATZ, D. R. Differences in accessory cell function. Immunobiology 168(1984)134-140.
5. LENZINI, L., ROTTOLI, P. and ROTTOLI, L. The spectrum of human tuberculosis. Clin. Exp. Immunol. 27(1977)230-237.
6. MERIGAN, T. C. A plaque inhibition assay for human interferon employing human neonate skin fibroblast monolayers and bovine vesicular stomatitis virus. In: In Vitro Methods in Cell Mediated Immunity. Bloom, B. R. and Glade, P. R., eds. New York: Academic Press, 1971, pp. 489-497.
7. RIDLEY, D. S. and JOPLING, W. H. Classification of leprosy according to immunity; a five-group system. Int. J. Lepr. 34(1966)251-273.
8. SHEPARD, C. C. WALTER, L., VAN LANDINGHAM, R. M. and YE, S. Z. Sensitization or tolerance to Mycobacterium leprae antigen by route of infection. Infect. Immun. 38 (1982) 673-680.
9. SHROFF, K. E., SAINIS, K. B., SENGUPTA, S. R. and KAMAT. R. S. Variation in immunogenicity of mycobacteria: role of antigen-presenting cells. Int. J. Lepr. 58(1990)58-64.
10. SHROFF, K. E., SENGUPTA, S. R. and KAMAT, R. S. Pathogenesis of route-related variation in T-suppressor response on immunization with mycobacterium. Int. J. Lepr. 58(1990)50-57.
11. SNYDER, D. S., Lu, C. Y. and UNANUE, E. R. Control of macrophage la expression in neonatal mice;role of splenic suppressor cell. J. Immunol. 128(1982)1458-1465.
12. STANFORD, J. L., ROOK, G. A. W., CONVIT, J., GODAL, T., KRONVALL. G., REES, R. J. W. and WALSH. G. P. Preliminary taxonomic studies on the leprosy bacillus. Br. J. Exp. Pathol. 56(1975)579-585.
13. STEEG, R. S., MOORE, R. N., JOHNSON, H. M. and OPPENHEIM. J. J. Regulation of murine macrophage la antigen expression by a lymphokine with interferon activity. J. Exp. Med. 156(1982)1780-1786.
14. VAN EDEN, W. and DE VRIES, R. R. P. Occasional review-HLA and leprosy: a re-evaluation. Lepr. Rev. 55(1984)89-104.
15. VAN EDEN, W.. ELFERINK. B. G., DE VRIES, R. R. P., LEIKER. D. L. and VAN ROOD, J. J. Low T-lymphocyte responsiveness to M. leprae antigen in association with HLA-DR3. Clin. Exp. Immunol. 55(1984)140-148.
16. WATSON, S. R., MORRISON, N. E. and COLLINS, F. M. Delayed hypersensitivity response in mice and guinea pigs to Mycobacterium leprae, Mycobacterium vaccae, and Mycobacterium nonchromo-genicum cytoplasmic proteins. Infect. Immun. 25(1979)229-236.
1. M.Sc, Research Fellow, Department of Immunology, Haffkine Institute, Parel, Bombay 400012, India.
2. M.D., Director, Department of Immunology, Haffkine Institute, Parel, Bombay 400012, India.
3. M.D., Assistant Director and Head, Department of Immunology, Haffkine Institute, Parel, Bombay 400012, India.
Reprint requests to Dr. Kamat.
Received for publication on 31 March 1989.
Accepted for publication in revised form on 17 August 1989.