- Volume 67 , Number 1
- Page: 46–51
Susceptibility of " et ," the spontaneously mutating CD1-derived nude mouse, to infection of M. lepraemurium
ABSTRACT
We have studied the susceptibility to infection by Mycobacterium lepraemurium (MLM) of a nude, hypothymic, CD 1-derived, spontaneous mouse mutant called " et " because of its extraterrestrial appearance. We found that despite their hypothymia, et/et mice were not more susceptible to infection by MLM than their euthymic et /+ counterparts. Infection of both et/et and et /+ mice with 50 x 106 bacilli by the intraperitoneal route led only to a mild infection with low levels of antimycobacterial antibodies and a small number of lesions. These lesions were indicative of reactive hepatitis and hyaline perisplenitis with lymphoid hyperplasia. Some small bacilliferous granulomas were also observed at the end of the experiment (5 months of infection). CD1 mice behave in a rather "resistant" manner to the infection by MLM. It is clear that the nu gene is not necessarily linked to the thymus defect, and it is also clear that the hypothymia of et/et mice does not obviously affect their general cell-mediated immune competence.RÉSUMÉ
Nous avons étudié la susceptibilité à l'infection par Mycobacterium lepraemurium (MLM), d'une souris nue, hypothymique, mutante spontanée dérivée de la souche CD-1, appelée " et " à cause de son aspect d'extraterrestre. Nous avons constaté que, en dépit de leur absence de développement thymique, les souris et/et n'étaient pas plus sensibles à l'infection par MLM que leurs homologues cuthymiques et /+. L'infection des souris et/et et et /+ avec 50 millions de bacilles par voie intra-péritonéale n'a produit qu'une infection légère avec des niveaux bas d'anticorps anti-mycobactériens et un nombre limité de lésions. Ces lésions étaient constituées d'hépatites réactionnelles et de périsplénites hyalines avec hyperplasie Iymphoïde. Quelques petits granulomes contenant des bacilles furent aussi abservés à la fin de l'étude (5 mois après l'infection). Les souris CD1 se comportent de façon plutôt "résistante" à l'infection par MLM. Il est clair que le gène nu n'est pas forcément lié à un déficit fonctionnel thymique, et il est également clair que le statut hypothymique des souris et/et n'affecte pas, de façon évidente, leur capacité globale à monter une réaction immunitaire à médiation cellulaire.RESUMEN
Estudiamos la susceptibilidad de los ratones " et " (llamados así por su apariencia extraterrestre) a la infección por Mycobacterium lepraemurium (MLM). Los ratones et/et son una mutante espontânea de los ratones CD1 que muestran hipotimia y alopecia (son ratones desnudos). Encontramos que los ratons et/et no son más susceptible a la infección por MLM que sus progenitores et /+, eutímicos y con pelo. Tanto en los ratones et/et como en los et /+, la inyección intraperitoneal de 50 x 106 bacilos produjo sólo una infección moderada con niveles bajos de anticucrpos anti-micobacterianos y poças lesiones en el hígado y en el bazo. Las lesiones l'ueron compatibles con hepatitis reactiva y con esplenitis hialina con hiperplasia linfoide. También hubieron algunos granulomas baciliferos al final del experimento (quinto mes de infección). Los ratones CD1 se comportait como los ratones de otras cepas "resistentes" a MLM. Es claro que el gene nu no está necesariamente ligado al defecto timico y también es claro que la hipotimia de los ratones et/et no afecta sensiblemente su competencia inntune celular general.In 1988, Rosas, et al . described the emergence of a nude mutant mouse derived from CD1 mice (11). Due to its extraterrestrial appearance this spontaneous mutant was called " et " (Fig. 1). Visual examination of the skin of this mutant revealed that some of the animals aged 40 days or older (but none younger) had patches of sparsely growing hair on their heads and limbs. A similar finding was observed by Flanagan (2 ) for nude ( nu/nu ) mice which were more than 10 days old. In addition, nude ( nu/nu ) mice were found to be without a thymus (7 ).

Fig.1. General physical appearance of the dorsal and ventral areas of both hairy parental(et/+) and nude (et/el) mice. Animals were anesthetized with chloroform and then bled by heart puncture through an opening in the skin as the ones shown.
On autopsy, Rosas, et al . found that et/et males have a degenerated thymus; whereas female et/et mice have a lymph node-like structure occupying the position normally occupied by the thymus. As expected, the atrophic male thymus and the female lymph node-like structure were lighter in weight than the eutrophic thymus of carrier ( et /+) and normal (+/+) CD1 mice (11).
Et/et mice were found to resemble nu/nu mice such that when reared together with their hairy siblings they were found to exhibit high rates of mortality during the first 2 weeks of life. However, when reared separately their survival time increased to 80% at 4 weeks and 60% at 52 weeks (11). Nude nu/nu mice when reared together with their hairy siblings showed a mortality rate of 55% within the first 3 weeks and of 100% at 25 weeks (2).
The majority of nu/nu mice die of general body weakness within 2 weeks of birth. The survivors grow slowly and may live for a considerable period of time, but usually all of the mice have died between 3 and 14 weeks of age (2). The short half-life of the nu/nu mouse is at least partially due to the absence of a thymus in these animals. The presence of a thymus is required for cellmediated immunity, and the absence of this organ therefore renders these animals highly susceptible to infection by a variety of microorganisms, in particular intracellular microorganisms such as Mycobacterium leprae and M. lepraemurium . The nu/nu mouse maintains the multiplication of M. leprae to levels close to 1010 bacilli per gram of tissue and can be inoculated with doses as high as 108 bacilli per foot pad without provoking an immune response, which would normally control multiplication of the microorganism (6). In contrast, normal mice of almost any euthymic strain will only allow proliferation of the microorganism for a limited period of time during which the disease is controlled and will eventually disappear (12). Nevertheless, it has also been shown that nu/nu mice infected with M. lepraemurium are capable of killing the microorganism following its early proliferation. This observation thus indicates that killing of the bacillus is not exclusively dependent upon T-cell activity (10).
In the present study we have compared the susceptibility of et/et mice with that of et /+ mice to infection by M. lepraemurium . The working hypothesis was that the relative hypothymia of et/et mice would cause these animals to be more susceptible to infection by M. lepraemtirium than animals of the parental euthymic ( et /+) CD1 strain.
MATERIALS AND METHODS
Infection and sampling
This study was carried out with groups of 10 et /+ mice and 10 et/et mice. All animals were males and were 2 months old at the start of the study. The bacilli ( M. lepraemurium , Hawaiian strain) were grown in NIH mice and were purified from infected tissue by using a combined methodology consisting of the entire procedure of Prabhakaran, et al . (9) and the Percoll step of Draper (1).
The mice were inoculated with 20 x 106 M. lepraemtirium by the intraperitoneal route. Progress of infection was monitored at monthly intervals for 5 months by taking blood samples (approximately 0.3 ml) from a small cut in the lateral tail vein. Sera were then used to look for antibodies to lipid and protein antigens of M. lepraemurium . Because the levels of anti-mycobacterial antibodies are found to increase in a manner parallel to the progression of infection, we considered the antibody titers to be a reliable although not an exact index of infection. At the end of the experiment (5 months of infection) three animals from each group were sacrificed and their spleens and livers removed for histopathological assessment of infection.
Histopathology and bacilloscopic studies
Tissue fragments taken from the livers and spleens of sacrificed animals were embedded in Tissue-Tek 4583 (Miles Inc., Elkhart, Indiana, U.S.A.). They were then frozen on dry ice and sliced into 6.0 µm sections in a freezing microtome (Tissue- Tek II, Cryostat; Miles). Sections were then processed and stained with hematoxylin and eosin (H&E) for general histology and with phenol fuchsin (Ziehl-Neelsen technique) to identify acid-fast bacilli (AFB).
Preparation of protein antigens
Protein antigens of M. lepraemurium were extracted by sonication of a 20% (v/v) suspension of the microorganism. The suspension was subjected to 15 1-min pulses at 80 watts in a sonicator (Sonics & Materials Inc., Danbury, Connecticut, U.S.A.) in the presence of 1 mM p-sulfonyl fluoride and 1 mM N-ethylmaleimide (both from Sigma Chemical Co., St. Louis, Missouri, U.S.A.). The sonicate was then collected and centriluged at 15,000 x g x 15 min. The resultant supernatant was recovered, adjusted to contain 1.0 mg of protein per ml and sterilized through Millipore membranes (0.22 j.im). Aliquots were then stored until further use at -20°C.
Preparation of lipid antigens
Whole lipids of M. lepraemurium were extracted following the method of Folch, et al . (3). Details on lipid extraction have been previously reported (5). The waxy material obtained after evaporation of the solvent was kept frozen at -20°C until further use.
Antibodies to protein and lipid antigens
Anti-mycobacterial antibodies present in the sera of infected animals were measured by using conventional enzyme-linked immunosorbent assays (ELISAs), as follows: a) wells (Maxisorp plates; Nunc, Napierville, Illinois, U.S.A.) were coated with 2.0 µg of protein [in 100 µl of borate buffer solution (BBS) pH 8.41 or with 5.0 µg of mycobacterial lipids in 100 µl ethanol; b) wells were washed four times with 150 µl of phosphate buffer solution (PBS) pH 7.4; c) blocking was carried out with 150 µl of 2% skim milk in BBS for 30 min; d) plates were incubated with 100 µl of the sera diluted 1:100 in 2% skim milk in PBS (60 min/37°C); e) wells were washed three times with 150µl (al of PBS; f) plates were incubated with 100 µl of horseradish peroxidase-labeled rabbit antibody to mouse immunoglobulins (1:2000 in 2% skim milk in PBS for 60 min at 37°C; g) wells were then washed three times with 150 µl of PBS; h) a mixture of 100 µl containing 4.0 mg of o-phenylenediamine (P1526; Sigma) and 10 Hi of 30% hydrogen peroxide (HI009; Sigma) in 10 ml of citrate buffer pH 5.0 was added to the wells; i) followed by 20 µl of 4 N sulfuric acid to arrest the reaction and, finally; j) absorbencies were read at 492 nm in an ELISA reader (Multiskan Plus Labsystems, Research Triangle Park, North Carolina, U.S.A.). BBS contains 0.3092 g of boric acid and 0.4768 g of sodium tetraborate per liter of 0.15 M NaCl; phosphate-citrate buffer, pH 5.0, consists of 0.1 M citric acid (24.3 ml), 0.2 M disodium phosphate (25.7 ml) and water (50 ml).
RESULTS AND DISCUSSION
Histopathology
Liver. The changes in histopathology were similar for both et/et and et /+ mice. The overall alterations observed included signs of congestion and focal rupture of the limiting plates by chronic infiltrates of lymphocytes. Hepatocytes showed evidence of reactivity and regeneration as manifested by binucleation, karyomegaly and the presence of nucleoli. In most centrolobulillar regions there were groups of lymphocytes associated with the necrosis of perivascular hepatocytes. Hyperplasia of Kupffer cells was also observed as well as some granulomas containing epithelioid cells.
Spleen. Changes in the spleen were also found to be similar for both the et/et and et /+ mice. The most outstanding alterations observed were the intense hyperplasia of the white pulp, congestion and slight-to- mild hemorrhage. In addition, diffuse thickening of the capsule with deposition of fibrinoid material on the plates was observed. These data were suggestive of hyaline perisplenitis with diffuse lymphoid hyperplasia.
Liver-associated bacilli. In the interstitial and centrolobulillar zones of the liver small groups of bacilli and isolated bacilli were observed. In addition, small granulomas made up of epithelioid histiocytes with scarce, although heavily stained bacilli were observed. Some globi (isolated bacilliferous macrophages) and small groups of two to six bacilliferous macrophages were also observed. These observations were found to be similar for both the et/et and et /-1- mice (Fig. 2).
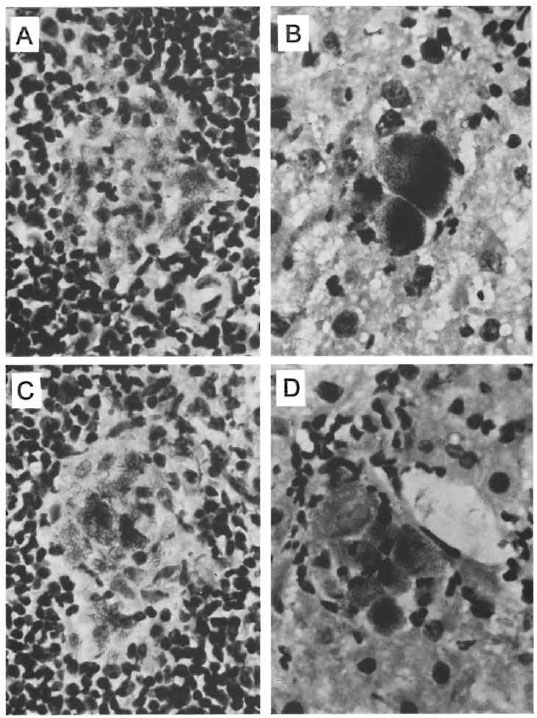
Fig. 2. Microscopic fields of spleen (left) and liver (right) sections taken from et/+ mice (A, B) and from ct/et mice (C, D). Fields were chosen and photomicrographs were taken from maximally developed lesions in each case (Ziehl-Neelsen x40).
Spleen-associated bacilli. Isolated bacilli and small groups of bacilli were observed in the interstitial spaces associated with regions of both the white and red splenic pulp as well as in the subcapsular zone of the spleen. The most affected region was found to be the white pulp where granulomas were more frequent than in other parts of the spleen. In spite of being scarce and small, these granulomas contained from low-to-moderate numbers of bacilli that were heavily stained. Similar changes in histopathology were observed in the spleens of both the et/et and et /+ mice (Fig. 2).
Antimycobacterial antibodies. Et/et and et /+ mice when infected with M. lepraemurium showed comparable curves of antibody produced against both protein and lipid antigens of the microorganism. Antimycobacterial antibodies were detectable from the first (for protein antigens) or the second (for lipid antigens) month of infection in both groups of animals. From the third to the fifth month of infection (when the study was stopped), the humoral response in the two animal groups was clearly evident although the response was not found to reach maximal levels in any of the mice assayed. Differences were observed between the et/et and et /+ mice; however these differences were not found to be statistically significant for any of the antigens or times used in this study (Figs. 3 and 4).
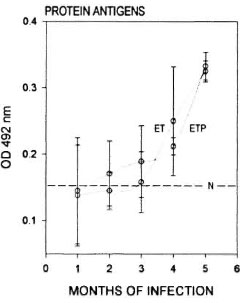
Fig. 3. Antibodies to protein antigens of M. lepraemurium from the sera of parental et/+ mice (ETP) and from et/et mice (ET). Readings were taken throughout the infection period with M. lepraemurium.
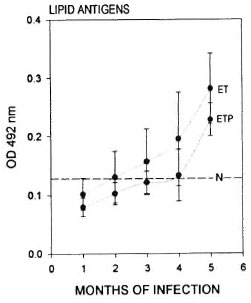
Fig. 4. Antibodies to lipid antigens of M. lepraemurium from the sera of parental et/+ mice (ETP) and from ellet mice (ET). Readings were taken throughout the infection period with M. lepraemurium.
Et/et mice represent a nude CD1 mutant that in addition to having a characteristic alopecia also possess a degenerated thymus, without being athymic. These animals are reared under conventional animal-house conditions, and they have a half-life of about 2 years. Although the immunological behavior of et/et mice has received little investigation, their thymic atrophy qualifies them as good candidates to be used as hosts in the study of susceptibility to infection by M. leprae and M. lepraemurium . The control of these mycobacterioses depends upon the host's ability to mount a competent cellmediated immune response (T-cell mediated). Our initial working hypothesis was that the relative hypothymia of the et/et mouse would render such animals more susceptible to infection by M. 'lepraemurium than the euthymic ( et /+) parental mouse. Our results have, however, indicated that this is not the case and that despite their hypothymia et/et mice were as susceptible (or resistant) to infection by M. lepraemurium as were the et /+ mice. This behavior clearly contrasts with that of the nu/nu mice which are truly athymic and, hence, highly susceptible to M. lepraemurium infection (4). It is clear that the morphologic defect in the thymus of the et/et mice does not correlate with an obvious functional defect in these animals, at least with regard to M. lepraemurium infection. The preliminary results of Marroquin, et al . (unpublished observations) have supported this finding, and have based their conclusions on the observations that both et/et and et /+ mice have: a) similar numbers of circulating leukocytes, b) comparable levels of CD4+ T cells, c) T cells that proliferate in a similar manner in response to concanavalin A, d) lymphocytes that produce similar amounts of IL-2 and, finally, e) similar primary responses to bovine serum albumin. Thus, nude et/et mice, although hypothymic, do not appear to have evident defects either in cell-mediated immunity or in their humoral immune reactivity. Other nude mutant mice that do not show alterations in the morphology or function of the thymus have also been reported (8).
Acknowledgment. This study was supported by the Dirección de Estúdios de Posgrado e Investigación del IPN (Project DEPI 970342) and by the Consejo Nacional de Ciência y Tecnologia (Project CONACYT 26427-M), México. The authors are fellow-holders from COFAA-IPN and/or the Sistema Nacional de Investigadores, SNI. México.
REFERENCES
1. Draper, P. Purification of Mycobacterium leprae . Report of the Fifth Meeting of the Scientific Working Group on the Immunology of Leprosy, Geneva, 24-26 June, 1980. Geneva: World Health Organization, 1980. (TDR/IMMLEP-SWG 5/80.3)
2. Flanagan, S. P. "Nude," a new hairless gene with pleiotropic effects in the mouse. Genet. Res. Camb. 8(1966)295-309.
3. Folch, J., Lees, M. and Sloane, S. G. H. A simple method for the isolation of total lipids from animal tissues. J. Biol. Chem. 226(1979)497-509.
4. Kawaguchi, Y., Matsuoka, M., Kawatsu, K., Homma, J. Y. and Abe, C. Susceptibility to murine leprosy bacilli of nude mice. Jpn. J. Exp. Med. 46(1976)167-180.
5. Luna, H. J., Rojas-Espinosa, O. and Estrada, P. S. Recognition of lipid antigens by sera of mice infected with Mycobacterium lepraemurium . Int. J. Lepr. 64(1996)299-305.
6. McDermott-Lancaster, R. D.. Ito, T, Kohsaka, K.. Guelpa-Lauras, C. C. and Grosset, J. H. Multiplication of Mycobacterium leprae in the nude mice and some applications of nude mice to experimental leprosy. Int. J. Lepr. 55(1987)889-895.
7. Pantelouris, E. M. Absence of thymus in a mouse mutant. Nature 217(1968)370-371.
8. Pantelouris, E. M. Nude mice with normal thymus. Nature 254(1975)140-141.
9. Prabhakaran, K.. Harris, E. B. and Kirch- heimer, W. F. Binding of 14 C-labeled DOPA by Mycobacterium leprae in vitro. Int. J. Lepr. 44(1976)58-64.
10. Resnick, M.. Bercovier, H.. Aizer, F., Mgr. N. and Levy, L. Death of Mycobacterium lepraemurium after multiplication in CBA, BALB/C and nude mice. Ann. Inst. Pasteur Microbiol. 138(1987)15-19.
11. Rosas, P., Castellanos, P. and Dominguez, R. The existence of a spontaneous hairless (nude) hypothymic mutant mice from the CD 1 strain, reared under conventional animal house conditions. Med. Sei. Res. 1(1987)553-555.
12. Shepard, C. C. The experimental disease that follows the injection of human leprosy bacilli into the foot pads of mice. J. Exp. Med. 112(1960) 445-454.
1. Sc.D.; Department of Immunology, Escuela Nacional de Ciências Biologicas, Instituto Politécnico, Carpio y Plan de Ayala, Colonia Santo Tomas, 11340 Mexico, D.F., Mexico.
2. B.Sc.; Department of Immunology, Escuela Nacional de Ciências Biologicas, Instituto Politécnico, Carpio y Plan de Ayala, Colonia Santo Tomas, 11340 Mexico, D.F., Mexico.
3. B.E., Department of Immunology, Escuela Nacional de Ciências Biologicas, Instituto Politécnico, Carpio y Plan de Ayala, Colonia Santo Tomas, 11340 Mexico, D.F., Mexico.
4. Sc.D., Department of Immunology, Facultad de Estúdios Superiores Zaragoza, UNAM, Mexico.
5. Sc.D., Department of Morphology, Escuela Nacional de Ciências Biologicas, Instituto Politécnico Nacional, Mexico.
Received for publication on 19 June 1998.
Accepted for publication in revised form on 3 December 1998.