- Volume 68 , Number 1
- Page: 11–7
The effect of ultraviolet light radiation on Mycobacterium leprae
ABSTRACT
Ultraviolet (UV) light is recognized as a potent sterilizing aid, but its relative effectiveness against Mycobacterium leprae has not been shown. We examined the influence of UV on the growth and metabolic activity of M. leprae harvested fresh f rom foot pads of nude mice. Temporary static suspensions were exposed to timed intervals of UV radiation generated f rom a fixed source to constitute dosages ranging f rom 0-12.64 x 104 erg/cm2. The metabolic activity of the bacilli was indexed by the oxidation of 1 14C-palmitate in BACTEC 12-B vials. The long-term effects of irradiation on cell division and growth were assessed by inoculation of BALB/c mouse foot pads. The metabolic activity in BACTEC showed an immediate dose-response-relatcd decline to a maximum of 50% of the control activity after exposure to 6.3 x 104 erg/cm2. Mouse foot pad studies showed a similar dose-response pattern. Effective-dose determinations based on metabolic or foot pad data wcrc similar. UV doses of 3.52 x 104 erg/cm2 resulted in an average 50% killing, and 7.73 x 104 erg/cm2 killed 84% of the M. leprae exposed. This UV sensitivity is similar to that reported for M. tuberculosis. UV sterilization and disinfection practices suitable for M. tuberculosis are likely to be equally effective for M. leprae.RÉSUMÉ
A lumière ultraviolette (UV) est reconnue comme un moyen puissant d'aide à la stérilisation. Cependant, son efficacité relative contre Mycobacterium leprae n'a pas encore été documentée. Nous avons examiné l'influence des UV sur la croissance et l'activité métabolique de M. leprae fraîchement récoltées des pattes de souris nues. Des suspensions temporairement statiques furent exposées à une irradiation UV de durée pré-déterminée, générée à partir d'une source fixe, constituant des doses de 0 à 12,64 x 104 erg/cm2. L'activité métabolique des bacilles fut indexée au moyen de la mesure de l'oxydation du "C-palmitate dans des flacons BACTEC 12-B. Les effets à long terme de l'irradiation UV sur la capacité du bacille à se diviser et croître furent évalués en inoculant des souris de souche BALB/c aux pattes. L'activité métabolique évaluée par le système BACTEC a montré une diminution immédiate dose-dépendante de celle-ci atteignant un maximum de 50% de l'activité contrôle après une exposition à 6.3 x 104 erg/cm2 Les études d'inoculation aux pattes de souris montrèrent une tendance similaire dose-dépendante. Les calculs de doses efficaces, basées sur les données métaboliques et de croissance dans les patles, furent également très similaires. Des doses d'UV de 3.52 x 104 et de 7.73 x 104 erg/cm2 ont provoqué une moyenne de 50% et une moyenne de 84%, respectivement, d'inactivation des Al leprae exposées. Cette sensiti vite aux UV est similaire à ce qui a déjà été rapporté pour Mycobacterium tuberculosis. Les pratiques de stérilisation et de désinfection aux UV utilisées pour M. tuberculosis sont probablement aussi efficaces chez M. leprae.RESUMEN
La luz ultravioleta (LUV) es un potente agente esterilizante, sin embargo no se ha establecido su electo sobre Mycobacterium leprae. En este trabajo examinamos la influencia de la LUV sobre el crecimiento y la actividad metabólica de Al leprae recién obtenido de la almohadilla plantar del ratón desnudo. Se expusieron suspensiones de Al leprae a diferentes dosis de irradiación UV durante diferentes periodos de tiempo. La irradiación UV fue emitida por una fuente lija a dosis que variaron de 0 a 12.64 x 104 erg/cm2. La actividad metabólica de los bacilos se midió en función de la oxidación del 14C-palmitato usando viales BACTEC 12-B. Los efectos a largo plazo de la LUV sobre la división y crecimiento de Al leprae se estudiaron por inoculación de ratones BALB/c en la almohadilla plantar. La irradiación con 6.3 x 104 erg/cm2 redujo rápidamente la actividad metabólica en BACTEC hasta un valor máximo del 50% con relación al valor observado en los bacilos no irradiados. Los estudios en la almohadilla plantar también mostraron un patrón similar de dosis-respuesta. Las determinaciones de las dosis efectivas por ambos métodos, resultaron igualmente similares. La dosis de LUV de 3.52 x 104 erg/cm2 provocó una mortandad promedio del 50%, mientras que 7.73 x 104 erg/cm2 mataron al 84% de las bacterias expuestas. La sensibilidad a la LUV de Al leprae es similar a la reportada para Al tuberculosis. Las prácticas de esterilización por LUV y de desinfección para Al tuberculosis, son probablemente igualmente efectivas para Al leprae.Exposure to ultraviolet (UV) radiation is one of the most commonplace disinfection procedures and best studied model systems for imparting DNA damage. Unfortunately, little information exists about the tolerance of Mycobacterium leprae to UV (6,12) Because of the increasing interest in genetic manipulation of M. leprae and the importance of adequate germicidal procedures as leprosy services are increasingly integrated into other clinical settings, we investigated the response of M. leprae to UV radiation.
Nearly all living organisms must somehow cope with the genotoxic effects of UV in their environment. Exposure to UV at wavelengths approaching the DNA adsorption maxima (~260 nm) causes pyrimidines to become covalently linked by the formation of four membered ring structures known as cyclobutan dipyrimidincs, or simply pyrimidine dimers. These dimers make for nonsense sequences or inhibit binding of proteins which may replicate or transcribe the DNA, support its structure, or be involved in regulatory feedback. The induction of such pyrimidine dimers is the classic test lesion for analysis of DNA repair and underlies the mechanism of action for UV disinfection (14). The energy dosage necessary to induce DNA injury varies across genera and is influenced by pigmentation, guanine cytosine (G-C) content and other variables (13).
The UV spectrum is subdivided into three wavelength bands designated as UV-A (400-320 nm), UV-B (320-290 nm), and UV-C (290-100 run). Solar UV consists mainly of UV-A and UV-B with UV-C wavelengths being mostly absorbed by ozone in the atmosphere. All UV spectra impart identical damage to DNA. However, lesions are more efficiently induced using the UV-C range (14). Therefore, we examined the response of M. leprae to varying dosages of UV-C using radiorespirometry to index its immediate effect on metabolic activity, and standard mouse foot pad techniques to discern its effect on growth in situ over the long term.
MATERIALS AND METHODS
UV irradiation. Cell suspensions were irradiated in a manner similar to the method of David (3). Specifically, a Gates MR-5 laboratory light (Bulbtronics, Farminsidalc, New York, U.S.A.) fitted with an F8T5GL low-pressure, mercury vapor germicidal bulb emitting maximum energy at 254 nm was used for UV-C irradiation. UV-C intensity levels were measured with a calibrated model 1400A International Light (IL) radiometer connected to an SEL 240 detector incorporating a diffuscr/filter combination that permits the system to read UV levels directly in units of microwatts per square centimeter (µW/cm2). The distance between the UV-C source and the organisms being irradiated was adjusted to deliver 150 µW/cm2 and was approximately 50 cm. The irradiances measured at the beginning and end of each experiment were averaged to establish the dose of UV-C delivered to the bacteria, and the averages never varied more than 4 µW/cm2 in any experiment. Each irradiancc interval was timed in seconds and total dosage delivered converted to erg/m2 for effective-dose calculations.
M. leprae. We harvested live M. leprae (isolate Thai-53) maintained in routine passage from the foot pads of BALB nu/nu nude mice (Harlan, Indianapolis, Indiana, U.S.A.) in a procedure published previously (1,4). The foot pads were cleansed in Bctadine®, and the skin removed before the highly bacilliferous tissue was excised and manually homogenized in 10 ml of 7H12 medium (Whcaton Science Products, Millville, New Jersey, U.S.A.). Excess tissue was removed by differential centrilegation, and after direct count enumeration, suspensions were diluted to contain I x 108 bacilli/ml in 7H12 medium. We exposed 500 µl of these suspensions in 10 x 35 mm polystyrene Petri dishes (Becton Dickinson Inc., Franklin Lakes, New Jersey, U.S.A.) to varying doses of ultraviolet light and evaluated its effect by radiorespiromctry and inoculation of the bacilli into foot pads of conventional mice for subsequent growth.
Radiorespirometry. We assessed the ability of each bacillary suspension to oxidize 14C-palmitate in BACTEC-12B media using a BACTEC 460 radiorcspiromctcr (Becton Dickinson) according to the method of Franzblau (4). An average growth index (Gl) was determined for each treatment dosage 7, 14 and 21 days after 1 x 107 bacilli were inoculated into triplicate vials and incubated at 33ºC. The cultures were held at 2.5% O2/10% CO2/Balance N2 (Air Liquidc America Corp., "Houston, Texas, U.S.A.), and this gas was replaced after each weekly reading. For effective-dose calculations with metabolic data, the mean GI resulting from the maximum radiation dose was considered the detection threshold and subtracted from the other mean GI readings before the test results were normalized as a percent of the untreated control activity and transformed to probits (13).
Mouse foot pad studies. To determine the effect of irradiation on the viability of M. leprae, we diluted the bacillary suspensions in Hanks balanced salt solution (HBSS; Gibco, Grand Island, New York, U.S.A.) and inoculated 30 µl aliquots containing 1 x 104 bacilli into both hind foot pads of conventional BALB/c mice (Harlan). Unirradiated control suspensions diluted 1:10 and 1:100 to reflect no treatment and -90% and -99% effective treatments also were used. Mice were harvested in a kinetic method (four from each group) at 30-day intervals after 90 days and when growth had been detected in all control groups similar to an earlier protocol (7). Growth was determined by direct enumeration of bacilli using a modification of the method of Shepard, et al. (10). Generally, the plantar surfaces of both hind feet were excised with scalpel and forceps and minced with sharp scissors before being transferred to a motorized Potter-Elvchjcm tissue grinder where it was homogenized to a fine paste for 1 min. Vcrsenc trypsin (I cc) in HBSS was added and homogenized with the tissue for an additional 30 sec before the entire unit was incubated for 15 min at 37ºC. After incubation the tissue was ground for an additional 30 sec and the contents were transferred to a glass Micklc homogenizcr with 25 glass beads, capped, and vibrated for 2 min.
For counting, 10 ul of the homogenized liquid was added to 10 µ1 of calf serum containing 2% phenol, mixed thoroughly and spread evenly over a 1 -cm2 area on a prcmarked counting slide. After drying in air, slides were fixed for 3 min in a covered staining dish containing 700 µ1 of formalin and then heated on a glass plate over a boiling water bath for 2 min. Hot slides were flooded and drained of distilled water containing 0.5% gelatin and 0.5% phenol twice, and then heated again for 2 min each time before being stained. Slides were stained using a modified Fite carbol-fuchsin method reported previously, and were decolorized for 30-40 sec with 5% sulfuric acid in 25% ethanol before counterstaining with methylene blue and a final wash (10).
Acid-fast bacilli (AFB) were enumerated by direct examination of 20 oil emersion fields along the vertical and horizontal axes of the stained smear using a calibrated microscope. The average number of bacilli per field was determined and multiplied by the appropriate calibration factor. The mean of three smears for each two foot pads was recorded and averaged with results obtained from the other individual mice in each group and postinfection interval. The means and standard deviations of direct bacillary counts from the foot pads were normalized to a percentage of the growth achieved by the untreated controls and compared to the theoretical -90% and -99% (103-102) control inoculations. These percentage data were then transformed to probits for effective-dose calculations (13).
RESULTS
Radiorespirometry. We determined the effective-dose range for UV injury to M, leprae in several preliminary studies using radiorespirometry. After exposing bacilli in shallow suspensions to varying doses of UV radiation, aliquots containing 1 x 107 M. leprae were inoculated into BACTEC I2B vials where their ability to oxidize 14C-palmitate in 7H12 medium at 33ºC was scored at 7, 14, and 21 days. BACTEC 460 respirometer readings ranged from 25 to 400 among all suspensions tested. Suspensions exposed to UV showed a dose-response related decline in respiratory activity detectable in the first 7 days of incubation when compared to control vials inoculated with unexposed suspensions.
The average metabolic activity determined from triplicate treatment sets was compared to the average for untreated controls and transformed to percentages. Figure 1 shows the response curve from respective runs. Values generally superimposed regardless of the initial metabolic activity of the stock bacterial suspension used. A decrease in metabolic activity was notable even after exposure to the lowest UV dosage tested. Exposure to UV above 12.49 erg/m2 appeared to have no meaningful effect on the metabolic activity of M. leprae. The maximum observed metabolic effect was only about 50% of the control activity in the first 7 days. All metabolic activity of UV-treated sets tended to wane somewhat over the ensuing 2 weeks of incubation but the weekly result did not change the shape of the dose response curve (data not shown). Accordingly, we focused our assessments on the activity measured at 7 days.
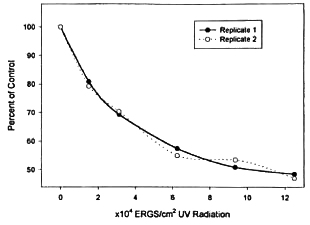
Fig. 1. Replicate dose response curves showing the percent decrease in metabolic activity detectable after 1 week in BACTEC-12B vials of M. leprae exposed to varying dosages of UV radiation as compared to the untreated control suspension receiving no radiation.
Mouse foot pad studies. To determine the effect of irradiation on the actual viability and growth of M. leprae, we inoculated aliquots of the suspensions into the foot pads of conventional BALB/c mice. The mice were inoculated into both hind foot pads with 1 x 104 bacilli in 30 ul of HBSS. Unirradiated control suspensions reflecting no treatment and control suspensions diluted to contain only 1x103 and 1 x 102 bacilli in 30 ul of HBSS were used to reflect growth at theoretical -90% and -99% rates. The mice were harvested in a kinetic method (four from each group) at 30-day intervals after 100 days of incubation, and the bacilli in each foot pad were enumerated and averaged (Fig. 2).
Fig. 2. Average and standard deviations of acid-l'ast bacilli detected in mouse foot pads between 100 and 190 days after inoculation with suspensions of 1x104 M. leprae which received varying dosages of ultraviolet radiation. Control suspensions receiving no (0 erg) radiation were inoculated at 1 x 104, 1 x 103 and 1 x 102 bacilli per foot pad to mimic results of 0% and -90% to -99% effective killing are shown in the first three groups.
The appearance of growth in the mouse foot pad and the timing of the plateau are relative to the number of viable bacilli in the inoculum (7). Growth was already approaching plateau level at >106 among the non-dilute untreated controls (containing 1 x 104 M. leprae) at 100 days, but it was below that level in each of the treatment groups and diluted control suspensions. The control samples diluted to reflect -90% and -99% growth of the untreated inoculum subsequently reached a plateau in later intervals. Growth among each UV treatment group also either reached a plateau or became detectable over the next 90 days. The timing of the plateau was delayed or the level of growth diminished among each correspondingly higher UV dosage group. Some growth was eventually detected in all of the UV treatment groups.
Growth in the mouse foot pads was detected in each untreated control during all intervals examined. When test samples showed growth in any 30-day interval which was between the level of the -99% and 100% untreated controls, the data were converted to a percentage of the growth seen in the undiluted untreated control. These percentage results ranged from 2% to 85%, and probit transformation was used to interpret the lethal-dose 50% (LD50), LD16 and LD84 in linear comparison to the log of the UV dose. The data were sufficient for effective-dose determinations in combination and at 100 and 130 days, and the percent effects interpreted were averaged. The averages and standard deviations found were: LD16% = 1.581 ± 0.049 x 104 erg/cm2; LD50% = 3.5213 ± 0.325 x 104 erg/cm2; and LD84% = 7.7378 ± 1.32 x 104 erg/cm2.
DISCUSSION
These data indicate that M. leprae are susceptible to UV injury and a dose of 3-7 x 104 erg/m2 is effective at killing 50%-90% of the organisms exposed. This level of sensitivity is similar to what we and others have observed for M. tuberculosis, but it is 2-5 times less than the UV sensitivity estimated for other common mycobacterial species such as M. smegmatis, M. kansasii, M. fortuituin and M. avium (2,3,5). These differences in sensitivity are considered relatively small (3). Other investigators have noted an apparent susceptibility of M. leprae to UV, but specific sensitivity ranges were not derived (6, l2). In comparison to other microorganisms the mycobacteria are fairly resistant to UV The range of sensitivity observed here ranks M. leprae similar to the Saccharomycetes and places them logarithmically more resistant to UV than Escherichia colt (11).
Among mycobacteria, differential sensitivity to U V is influenced by the size of the bacterial genome, the cellular concentration of carotenoids, and the efficiency of DNA repair systems. Species with long generation times theoretically have more opportunity to effect DNA repair and may thus appear to be more resistant to UV. M. tuberculosis is suggested to have a relatively inefficient DNA repair system because it is somewhat more sensitive to UV than other mycobacteria but has a rather long 2.5 clay generation time (3). M. leprae has a generation time of 12.5 days (7). The high sensitivity to UV seen here for M. leprae would suggest that it is at least as equally inefficient at DNA repair as M. tuberculosis. The reasons underlying these inefficiencies are unknown, but species which live primarily in an intracellular environment may have little need for efficient DNA repair systems.
These studies were done with suspensions containing a high percentage of viable bacilli derived from fresh nude mouse tissues. Death by UV injury is the result of an accumulation of pyrimidene dimcrs in the DNA (2). Sensitivity to UV injury is not influenced by the percent viability or metabolic state of the organisms tested. Therefore, suspensions from tissues of other hosts that may have lower percent bacterial viability or metabolic activity would be expected to show similar sensitivity to UV.
M. leprae show no significant cell division in vitro, and the response to UV injury measured by metabolic activity was not directly proportional to the effect seen in vivo. Although mouse foot pad studies clearly showed greater than 99% killing in some suspensions tested, the maximum reduction in the metabolic activity noted was only to about half that of the untreated control. These results are in keeping with similar observations using readiorespirometry for in vitro drug-susceptibility assays, and indicate that some aspects of metabolism and active transcription continue in vitro which were disrupted either directly or indirectly by UV exposure (4). In addition, if the extremes of the metabolic activity range are considered as 0% and 100% effects, then the ED50% dose determined for M. leprae would be similar whether it was calculated from in vivo or in vitro data. With such limitations in mind, oxidation of palmitate may be useful as a rapid objective measure of viability for M. leprae.
Growth of M. leprae in the mouse foot pad is censored on one end at a plateau caused by the induction of the host's cellmediated immunity, and limited on the other end by the sensitivity of the detection method. Levy (7) has shown that the time to appearance of bacterial growth and its plateau in the mouse foot pad is a function of the proportion of organisms in the inoculum which are capable of multiplying. Relatively few viable bacilli are required for growth to reach plateau levels in the mouse foot pad, the main difference being the fewer number of viable organisms present in an inoculum the longer the time required for growth to become detectable or reach a plateau. The offset in time to growth detection and plateau seen here among the UVtreatcd sets and diluted control suspensions results from a decrease in the number of viable organisms in those inocula. Some growth was eventually detected in each of the UV exposure dosages tested, and with cell clumping and other physical barriers it is difficult to obtain complete sterilization with UV radiation.
UV lamps have been used effectively for many years to aid infection control of aerosolized pathogenic organisms, disinfecting the air or laboratory surfaces. Their utility has been reviewed in depth by Riley and Nardcll (9). Health professionals should remember that UV radiation is hazardous and overexposure can result in acute eye or skin injuries for their personnel (8). The sensitivity to UV seen here for M. leprae suggests that containment and disinfection practices generally found suitable for M. tuberculosis should be equally effective against M. leprae.
Acknowledgment. We are grateful for the technical assistance given by Rita Sanchez, Roena Domingue and J. P. Pasqua. These studies were supported in part with funds from the National Institute of Allergy and Infectious Disease (contract number AI5015-01).
REFERENCES
1. Adams, L. B., Gillis, T. P., Hwang , D. II. and KRAHENBUHL, J. L. Effects of essential fatty acid deficiency on prostoglandin E2 production and cell-mediated immunity in a mouse model of leprosy. Infect. Immun. 65 (1997) 1152-1157.
2. David, H, L., JONES, W. D. Jr. and NEWMAN, C. M. Ultraviolet light inactivation and pholoreactivation in the mycobacteria. Infect. Immun. 4 (1971)318-319.
3. David, J. W. Response of mycobacteria to UV light radiation. Am. Rev. Respir. Dis. 18 0 (1973) 1175-1185.
4. FRANZBLAU. S. G. Drug susceptibility testing of M. leprae in BACTEC 460 system. Antimicrob. Agents Chemother. 33 (1989) 2115-2117.
5. Huber, T. W., Reddick, R. A. and KUBICA, G. P. Germicidal effect of ultraviolet irradiation on paper contaminated with mycobacteria. Appl. Microbiol. 19(1970) 383-384.
6. Kaur, S., Kumar, B., Ganguly, N. K. and Ciiakravarii, R. N. Viability of M. leprae under normal and adverse atmospheric conditions. Lepr. India 54 (1982) 228-233.
7. Levy, L. Death of Mycobacterium leprae in mice and the additional effect of dapsone administration. Proc. Soc. Exp. Biol. Med. 135 (1970) 745-749.
8. Murray , W. E. Ultraviolet radiation exposures in a mycobacteriology laboratory. Health Phys. 58 (1990)507-510.
9. RlLEY, R. L. and Nardell, E. A. Cleaning the air: the theory and application of ultraviolet disinfection. Am. Rev. Respir. Dis. 139(1989) 1286-1294.
10. Shepard, C. C. and McCRAE, D. H. A method for counting acid-fast bacteria. Int. J. Lepr. 36 (1968) 78-82.
11. Spector, W. S. Handbook of Biological Data. Philadelphia: W. B. Saunders, 1956, p. 474.
12. SREEVASTA, K. K. Viability of M. leprae while undergoing laboratory procedures. Indian J. Lepr. 69 (1997) 353-359.
13. Tallarida, R. J. and Murray, R. B. Manual of Pharmacologic Calculations with Computer Programs. 1st. edn. New York: Springer-Verlag, 1981, pp. 19-21.
14. WALLACE, S. S., va n HOUTEN, W. and Kow, Y. W. Environmental damage to DNA. In: Annals of the New York academy of Sciences, DNA Damage: Effects on DNA Structure and Protein Recognition. 1st. edn. New York: New York Academy of Sciences, 1994, pp. 24-28.
1. Ph.D., Hansen's Disease Center at Louisiana State University, P.O. Box 25072, Baton Rouge, LA 70894, U.S.A.
2. Ph.D., GWL Hansen's Disease Center at Louisiana State University, P.O. Box 25072, Baton Rouge, LA 70894, U.S.A.
Reprint requests to Dr. Truman at the above address or FAX 1-225-346-5786; e-mail: rtrumal@lsu.edu
Received for publication on 1 October 1999.
Accepted for publication 23 November 1999.